
The Design of Giardia and the Genesis of Giardiasis
Abstract
Giardia is a genus of protozoa discovered in 1681. Six morphologically distinct species are recognized. It mainly attaches in the upper GI tract of a wide variety of vertebrates (including zebrafish), often with beaver and muskrat as reservoirs/carriers but exhibiting minimal—if any—disease in some animals. Giardia is usually non-pathogenic in the human population, even in children if exposed early in life. Although Giardia can be pathogenic, some strains colonize the gut with no malady. This parasite is not invasive and only serious infections depress the small intestine. Giardia are pear-shaped, have an adhesive disc for attaching to enterocyte cells in the small intestine villus, and move with eight designed flagella. In the post-Fallen world, Giardia infection occasionally has resulted in digestive dysfunction. However, Giardia may function in non-parasitic, possibly mutualistic, ways. For example, it may have been designed to aid digestion having a role as a “primer.” The presence of Giardia muris causes a fundamental change in the microbiome in mice and Giardia may have other influences on the microbiome such as enhancing digestion in certain animals and possibly shifting ratios of bacteria from anaerobic to aerobic. Giardia may play a role in host metabolism and provide nutritional enhancement via its association with enteric bacteria, like E. coli. The function of Giardia may parallel with non-parasitic tasks found in Trypanosoma lewisi, and also termite systems that contain protozoa and bacteria for plant digestion. Giardia has two “faces” even in today's world: a harmless commensal in wildlife and a pathogenic parasite in humans.
Keywords: origin of Giardia, pre-fall, post-fall, Giardia design, genesis of giardiasis (beaver fever), parasite origins; microbiota design; microbial digestion, immune stimulator
Introduction
Giardia is a unique, if not attractive, organism and has long been a favorite of microscopists (Fig. 1). It has appeared on the cover of several microbiology and parasitology texts in recent years. Microbiology students when viewing Giardia through the microscope have been known to exclaim, “He’s cute!” Well, Giardia may look cute through the microscope, but Giardia will not feel cute if they over-colonize the intestine. Giardia has been described as a “cartoon,” a monkey face, smiling face, etc. It is photogenic, pathogenic, and has a portentous personality all its own (Table 1). The reason that Giardia looks like a face is its cell features: nuclei (its “eyes”), median bodies (its “mouth”), axonemes (its “nose”), and flagella (its “hair”) (Figs. 1 and 3).
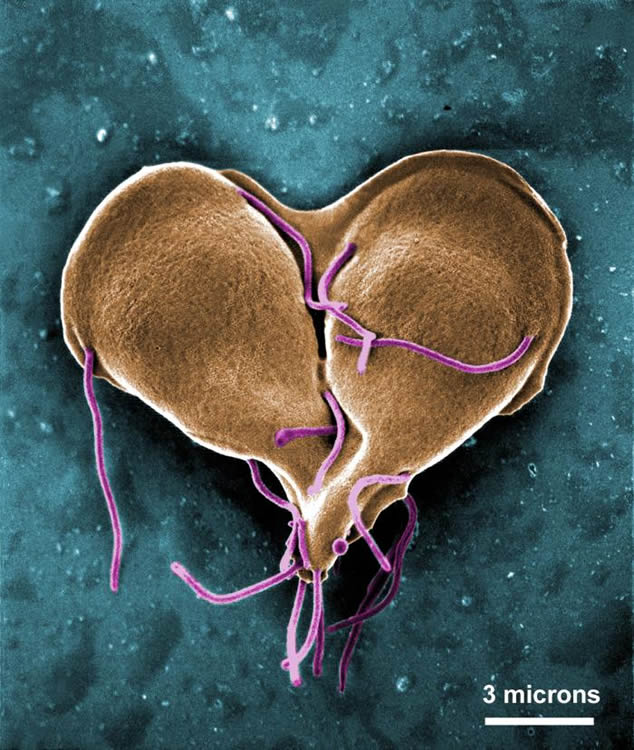
Figure 1. Giardia lamblia dividing. (A false-color scanning electron micrograph of trophozoites undergoing mitosis. Image courtesy of Centers for Disease Control and Prevention.
Table 1. Giardia at a Glance
- Giardia is a genus of protozoa found in the GI tract of vertebrates, often with beaver and muskrat as reservoirs/carriers but exhibiting minimal— if any—disease in some animals, like ducks and geese.
- Giardia are pear-shaped, have an adhesive disc for attaching to the small intestine, and move by eight flagella.
- Although this parasite can be pathogenic (different strains produce different pathogenicity), some colonizations reveal minimal malady in beavers and muskrats.
- In the post-Fallen world Giardia infection has resulted in digestive dysfunction, disease, and diarrhea in dogs, cats, and humans.
Giardia (in humans known as Giardia intestinalis, Giardia lamblia, or Giardia duodenalis) is found in or on soil, food, or water that has been contaminated with feces from infected humans or animals. There are about 40 species that have been described from various animals, but many of them are now considered the same species (or subspecies). Creation biologists consider them the same kind or baramin. In standard texts, six morphologically distinct species are recognized. Giardia infects humans and other mammals (especially beavers). G. muris is found in mice and rodents (Roberts, et al. 2013). Other Giardia species are found in voles, birds, amphibians, and even zebrafish. In many wild animals the parasite is not pathogenic. In some beaver populations, Giardia appears to stimulate the immune response early, so later in life the beavers have protection against other pathogens and parasites.
Purpose/Objectives of Paper
Giardia has long been an interest to microbiologists, parasitologists, epidemiologists, and creation and evolutionary biologists. Giardia, and the resultant disease, giardiasis, is notorious. Yet, we read in Genesis 1:31 that God made everything “very good.” If everything that God made was good, where did Giardia come from? Was Giardia created along with the rest of life in the first week of Creation, or are they a result of the Curse? Were there protozoans residing in the GI tract of animals and man before the Fall? These and other questions have been asked by creation biologists (Gillen 2014), and their answers may surprise you.
The objectives of this paper attempt to answer:
- Why did God create Giardia?
- In a post-Fallen world, how is Giardia pathogenic?
- How have Christians been involved in key discoveries linking Giardia to diseases?
The Anatomy of Giardia
Giardia intestinalis is a binucleate, flagellated protozoan with a life cycle that alternates between an actively swimming trophozoite and an infective, resistant cyst (Fig. 2). Trophozoites measure 8–18 μm in length, from their broad (anterior) end to their narrow (caudal or posterior) end, up to 10 μm in breadth (Fig. 3). The trophozoite shape resembles that of a pear cut in half along it long axis (a pyriform shape). On the anterior half of the flattened ventral surface is located a distinctive concave disc with a raised anterior ridge at the end. This ventral disc is used to attach the trophozoite to the epithelium of host enterocytes (Fig. 4), so that the parasite is not swept away with digested food. If the attachment is lost, the trophozoite passes out of the body and is unable to survive. Attachment onto the surface of the enterocyte is therefore crucial. The microscopic sucker holds the trophozoite fast to its host, and the outline of the ventral disc can be seen impressed on the surface of affected enterocytes (Marquardt, et. al. 2000).

Figure 2. Giardia Life cycle. The life cycle of Giardia lamblia, as described by the Centers for Disease Control and Prevention.
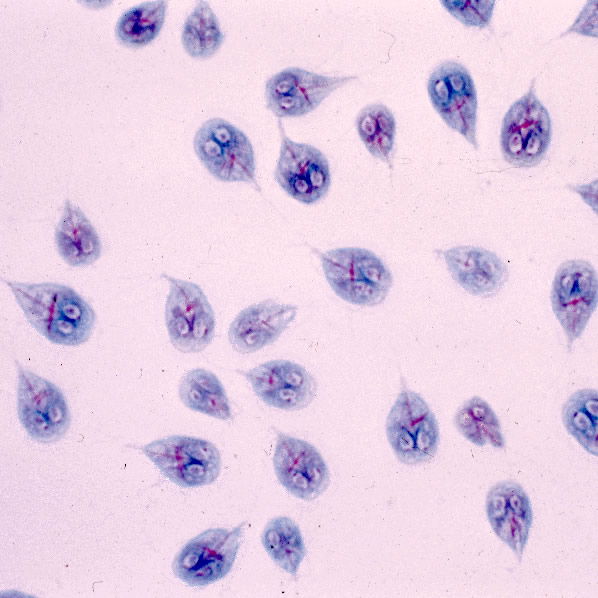
Figure 3. Giardia trophozoite. The trophozoite stage of Giardia lamblia has a face with two nuclei that look like eyes. Image from Centers for Disease Control and Prevention.
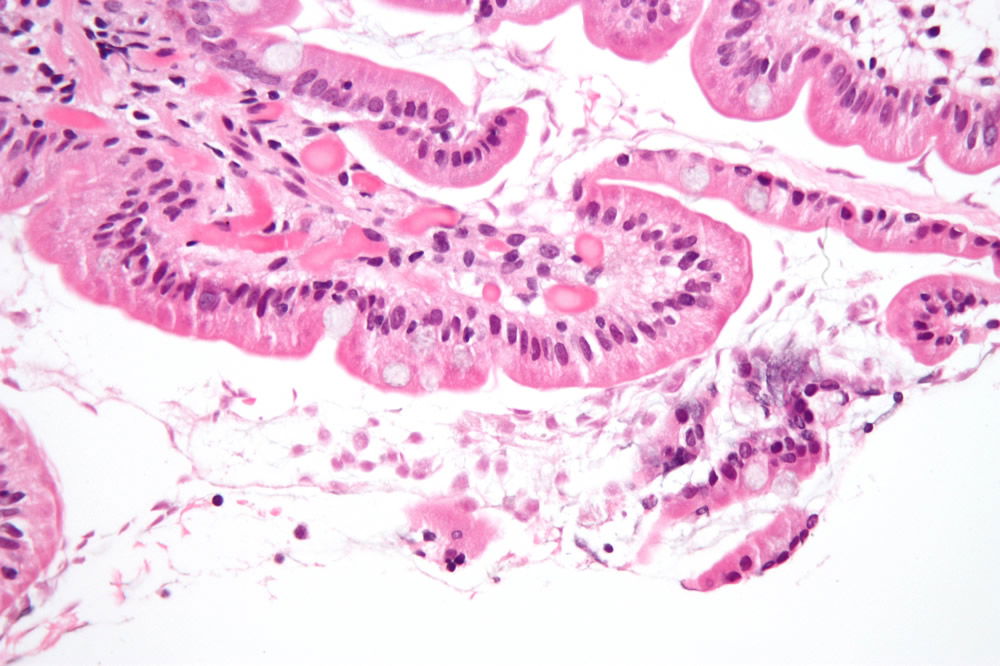
Figure 4. Giardiasis in the duodenum. Image by Nephron, via Wikimedia Commons.
History of Giardia
Many believe that Anton van Leeuwenhoek (a Dutch Reform Christian) was the first to describe Giardia lamblia (Gillen and Oliver 2012). He also founded advanced parasitology using his crude, homemade microscopes. Leeuwenhoek (Fig. 5) observed Giardia, sheep liver flukes, and many other microscopic creatures. In 1681, he examined his own stool samples during times of diarrhea. In his runny stool, he found “little animals.” Leeuwenhoek described what would later be termed the trophozoite stage of Giardia. He said it was a slow moving animal, but able to make quick motions with its “paws.” Today, we know this is a helical motion caused by the eight amazing flagella. It was not until 1880 that biologists realized that there were other stages in Giardia that did not contain flagella. The vegetative stage is now known as the cyst and is resistant to environmental factors, such as stomach acid and chlorine (Fig. 6). Today scientists can see molecular motors and other submicroscopic cellular machinery that would have utterly stunned Leeuwenhoek. He shared his hand-drawn illustrations of his discovery with Robert Hooke (also a famous Anglican Christian scientist) who verified them.

Figure 5. Leeuwenhoek painting. Leeuwenhoek discovered Giardia in 1681. Image by Fæ, via Wikimedia Commons.
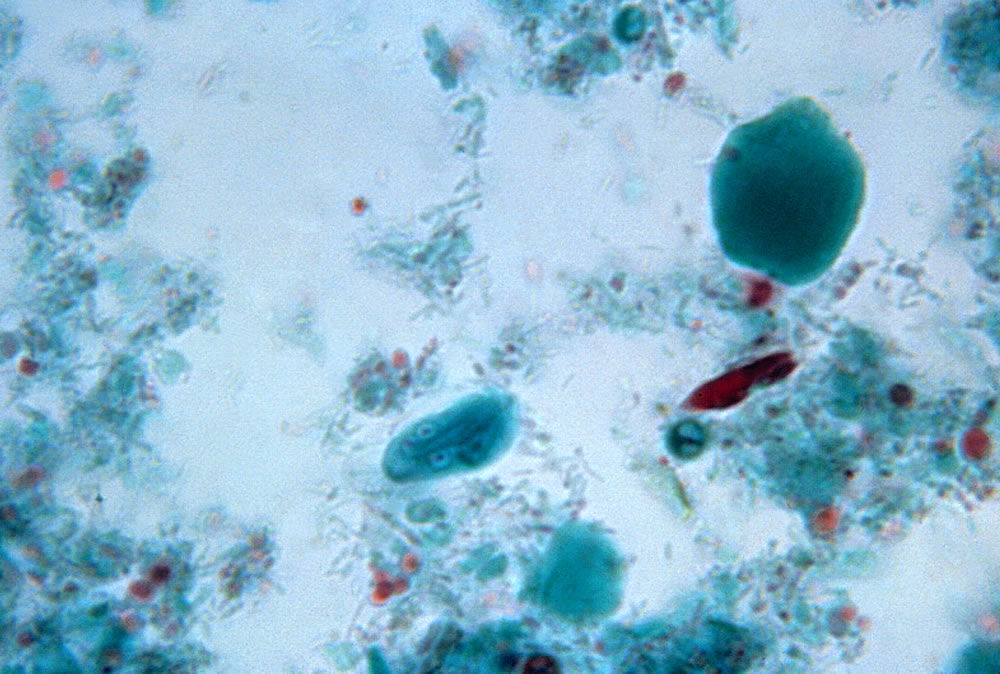
Figure 6. Giardia cyst. The cyst stage of Giardia lamblia has a face with four nuclei that look like eyes. Image from Centers for Disease Control and Prevention.
In 1681, Leeuwenhoek described Giardia and its movement this way:
All the particles aforesaid lay in a clear transparent medium, wherein I have sometimes also seen animalcules a-moving very prettily; some of ’em a bit bigger, others a bit less, than a blood globule, but all of one and the same make. Their bodies were somewhat longer than broad, and their belly was flatlike, furnisht with sundry little paws, where with they made such a stir in the clear medium and among the globules that you might e’en fancy you saw a pissabed [a pillbug that resembles Giardia] running up against a wall; and albeit they made a quick motion with their paws, yet for all that they made but slow progress. (Dobell 1932, p. 224)
The most significant and extensive biography of Leeuwenhoek was written by Dr. Cecil Clifford Dobell (1886–1949) of London, England (Fig. 7). Dr. Dobell was also the first to show in 1921 that Giardia was a pathogen. From the time of Leeuwenhoek (1681), many others enhanced and fine-tuned the description and life cycle of Giardia and a few even suggested that it might cause diarrhea. But, it was Dobell who proved Giardia was the cause of this unfortunate condition. He was an accomplished protozoologist and also studied algae and other intestinal amoebae besides Giardia. Like Leeuwenhoek and Hooke, it is believed that Dr. Dobell was a Christian (Anglican Church).

Figure 7. Clifford Dobell in 1949. Dr. Dobell first provided proof that Giardia is a pathogen. Image by Materialscientist, via Wikimedia Commons.
Protozoan Symbiosis
Protozoologists have found that a vast majority of flagellated protozoa are a valuable part of the ecosystem and do not cause pathology in man or animal. Flagellated protozoa that are parasitic (e.g. Giardia intestinalis) have closely related species that are harmless commensals such as Chilomastix mesnili. Even with the parasitic G. intestinalis there is a large number of people infected but exhibit no evidence of the disease. This is also true with the trypanosomes (other flagellated protozoans), some of which cause much suffering and death. Trypanosoma cruzi is the causative organism of the deadly Chagas’ disease of South America and Trypanosoma brucei is the causative agent of African sleeping sickness; however, past decades have shown a beneficial side of Trypanosoma (Table 2). For example, from 1950 to the 1970s, Dr. David Lincicome of Howard University studied Trypanosoma duttoni and T. lewisi. Lincicome and co-author Lee reported, “Weight gains in mice fed a pantothenic acid (Vitamin B5)-deficient diet and infected with [Trypanosoma] duttoni ranged up to 12% over uninfected controls and up to 41% in pair-fed controls” (Lincicome 1971).
Table 2. Evidence of Pre-Fall Benefits of Gut Protozoans
Work of Dr. David Lincicome (Howard University) 1950–70s
- Studied Trypanosoma duttoni and T. lewisi
- Found weight gain and improved health in rats & mice with B-vitamin deficiency (esp. Vitamin B-12) when colonized by Trypanosomes (Marquardt, et al. 2000)
- Concluded they co-evolved to bring more than just tolerance, but a benefit to both, akin to E. coli and mammals
- Creation view: glimpse of original design and purpose of flagellated protozoans; good design, not disease in a pre-Fall world.
- Original very good creation effaced by pathogens (T. brucei and T. cruzi), but not erased (T. duttoni and T. lewisi).
Additionally, research by Lincicome and students showed that food intake and body weight changes of rats inoculated with Trypanosoma showed significant increases over the noninoculated controls (Lincicome and Sheppersox 1963). Such research lends support to the hypothesis that the trypanosomes were mutualistic (or commensal) members of the “normal” microbiota in the pre-Fall world. In fact, the evolutionary researchers concluded that the trypanosomes studied co-evolved (via so-called convergent evolution), bringing more than just tolerance but also a benefit to both trypanosome and the “infected” mammal, much like certain strains of E. coli in mammal GI tracts today.
The fact that there are trypanosomes much less pathogenic than other strains (such as T. cruzi) could be an indication of emerging pathogenicity after the Fall, just thousands of years ago. Indeed, molecular evidence indicates humans may have been suffering from Chagas’ disease starting about 4000 years ago (Roberts, et al. 2013).
Giardia Symbiosis
As reported by wildlife biologists, Giardia colonizes beaver and muskrat GI tracts, but do not cause disease. Perhaps, in a pre-Fallen world, Giardia aided digestion of plant and wood matter due to its “priming” the creature’s microbiota. Experimentally, biologists (Barash et. al. 2017) know the presence of Giardia muris causes fundamental shifts of normal microbiota in mice (Fig. 8). Specifically, Giardia colonization is typified by microbial expansion of aerobic Phylum Proteobacteria (ex. E. coli) and decreases in anaerobic Phyla Firmicutes (ex. Clostridium) in mouse foregut and hindgut experiments. But, we know a shift in the ratios of families of bacteria that are anaerobic to different families that are more aerobic in mice suggest that they may provide nutritional enhancement with bacteria, like E. coli (Fig. 9). The function of Giardia may parallel with non-parasitic tasks found in Trypanosoma lewisi, and also with termite systems that contain protozoa and bacteria for cellulose digestion.

Figure 8. Giardia influence the normal microbiota. Illustration by Jayne Gillen.
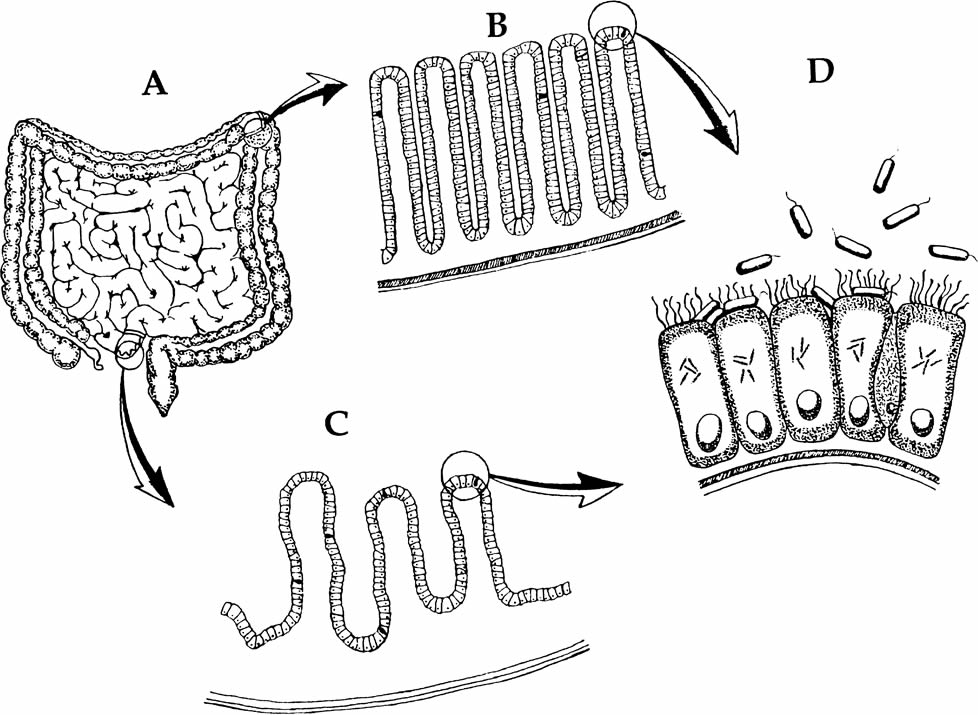
Figure 9. E. coli and Intestines. E. coli provides vitamins K and B-12 for the body due its colonization in the intestines. Image from Gillen, 2009, Body by Design, 58.
Experiments with mice demonstrate a reduction of the numbers of obligate anaerobic bacteria from the phylum Firmicutes (Barash et. al, 2017). That is, indirectly, Giardia may inhibit notable pathogenic species of this Phylum. These include: Clostridium perfringens (gangrene, food poisoning) and Clostridium difficile (CDIF). CDIF causes pseudomembranous colitis, a severe form of diarrhea. Giardia may serve as a protective agent against pathogens and toxins (ex. CDIF) in certain animals and in children (Barash et. al. 2017).
We might think of Giardia with its associated bacteria as a “Point Guard” in basketball, or perhaps the “master engineer.” The point guard is one of the five positions in a regulation basketball game. He has perhaps the most specialized role of any position. Point guards are expected to run the team’s offense by controlling the ball and making sure that it gets to the right players at the right time. The presence of Giardia in the small intestine may biochemically direct bacteria in the bowel to operate more effectively. Perhaps this may be accomplished via unique secretions from Giardia (e.g. alanine) that are specific for G-protein receptors located on bowel bacteria.
Other possible positive roles of Giardia include that of a glucose regulator. Giardia may also be a possible immune stimulator of IgA. Many children that have high IgA levels are colonized with Giardia and show no signs of infection or disease. Indeed, early exposure to Giardia in many children (Fig. 10) gives them “immunity” for life, serving as a type of “vaccine.” However, the disease state occurs with elevated numbers of Giardia on the villi, their displacement from original animals intended, and their modification from their original kind and loss of information (mitochondria / mitosome reduction). These are the two faces of Giardia: the pre-Fall functions of possibly aiding in digestion, stimulating the immune system, and in keeping a balanced gut microflora; and the post-Fall world, where they are causing dysfunction, disease, and diarrhea.

Figure 10. Giardia acquired by children drinking clear water from stream. Image by Fæ, via Wikimedia Commons.
Giardia Movement: an Intricate and Irreducibly Complex Design
There is certainly nothing simple regarding the sophisticated bacterial flagellum, but the Giardia flagellum story is even more complicated. In the article “Magnificent Motors: God Invented It First” (Purdom 2012), Dr. Purdom describes Giardia’s marvelous miniaturized motion (Fig. 11). The flagellum can move in a helical wave in addition to rotating “not just one, but four pairs of flagella (tentacle-like arms) extend from its body.” They enable Giardia to swim with a rotating motion, much like that of a beater on an electric mixer.
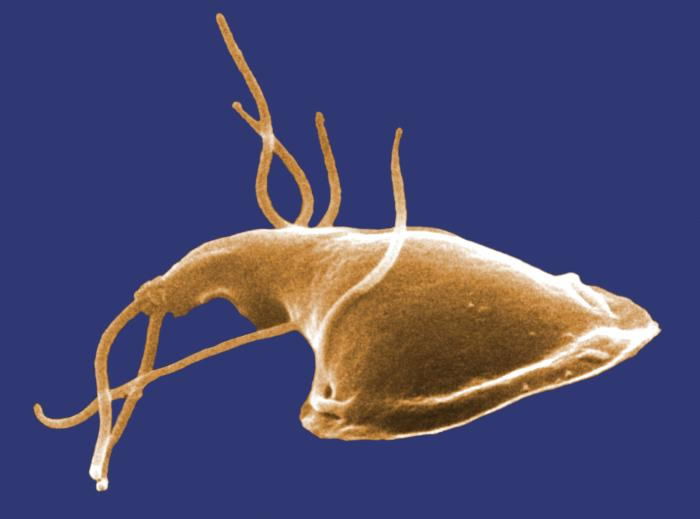
Figure 11. Magnificent Motors. Giardia has intricately designed flagella and their movement is like a personal submarine. Image by 7mike5000, via Wikimedia Commons.
Giardia is unusual in that it can infect many different types of hosts—both humans and animals. The key to its versatility is that each of the four pairs of flagella has different movements and functions, allowing it to infect very different types of hosts. In summary, the best analogy for Giardia motion is that of an active personal submarine. It can maneuver and hover to precise locations in the duodenum (Fig. 4).
The Complex Design of Helical Movement by Flagella
The “sundry little paws” that Leeuwenhoek speaks of refers to the flagella of Giardia (Gillen and Oliver 2012). Clearly they are incredibly complex. For example, just one flagellum possesses enormous intricacy, but Giardia possesses four pairs of flagella, which independently move in different patterns to increase the mobility of the organism. This requires much more complexity than just a “standard” bacterium with a single flagellum. This is a testament to the creative power of God who created this single-celled creature with its detailed function to show His glory.
The four pairs of flagellum each serve a different function. Two pairs act like paddles, one moves with a wave-like action akin to a snake moving through water, and the last pair runs along the entire length of the body and helps to increase the flexibility of the tail end (Purdom 2012). All of the pairs work in sync with one another to help the Giardia trophozoite move in unique ways so it can attach to its host properly. According to a study conducted by Lenaghan (2011), the free swimming motion of trophozoites revealed that when they were moving rapidly, the caudal region of the cells displayed a wave-like flexion. A possibility was that the flagella had a role in controlling the stability and directionality of the cell and directionally controlled the motion. The hypothesis, however, was that the flexion of the caudal region of the trophozoite allows for the increased speed associated with 3D swimming. In addition, previous studies have also shown that the internal portions of the caudal flagella were capable of movement; so combined with the lack of cytoskeletal components in the caudal region of the cell, the internal beating of the caudal flagella is responsible for the movement of the entire caudal region of the Giardia trophozoite (Lenaghan et al. 2011). It was also found that the caudal flagella are bound within the cytoplasm for almost the entire length of the cell, and the internal movement of these flagella would cause the cell to change shape as a result. This shows a great flagellar complexity, since the movement not only affects the flagella themselves, but also the entire trophozoite.
Most scientists would say that this amazing mechanism evolved by chance over millions of years, but could that be the case? If just a portion of the information that makes up one of the flagella was missing or out of place, the entire mechanism would fail and the organism would not survive. This concept is known as irreducible complexity, where a single system composed of one or more well matched, interacting parts contribute to the basic function of the organism. In addition, the absence of any one of these functioning parts causes the entire system to stop operating (Behe 2006).
Giardia “Evolution”
Giardia was once thought to be a breakthrough in evolutionary biology, possibly bridging the gap between prokaryotes and eukaryotes. Due to its “primitive” lack of mitochondria, it was thought Giardia diverged from one of the earliest lineages of eukaryotes before the alleged endosymbiotic relationship of mitochondria began (kingdom Archezoa). However, this evolutionary model came under revision since the discovery that virtually all Archezoa proved to have a set of mitochondrial genes in their nuclei. Meanwhile, sequencing the genome of G. intestinalis is complete, but results are not conclusive about its origin. So, Giardia was once thought to be a simple cell by evolutionary biologists due to their lacking mitochondria and was dubbed a cellular missing link. Today, evolutionary biologists understand that Giardia have mitosomes (mitochondria-like bodies) and have changed their view. They now believe a non-mitochondrial state is not a primitive trait, but is the result of reductive evolution. Creationists would call it devolution – a loss of genetic information.
The Genesis of Giardiasis
Giardia is a pathogenic protozoan in today’s world causing a gastrointestinal illness called giardiasis, characterized by severe diarrhea that can last several (4) weeks. Giardia is a distinguished flagellate in that it has four pairs of anterior flagella and two nuclei that stain darkly, giving the appearance of having eyes in its trophozoite (vegetative) stage. Giardia can be divided along its longitudinal axis and is therefore said to display bilateral symmetry. Once it starts dividing, considerable numbers can build up in the intestine (Fig. 12).

Figure 12. Giardia in high numbers. Giardia lamblia attached to the epithelium of the small intestine by a ventral adhesive disc, allowing the parasite to absorb nutrients from the lumen of the small intestine. Image by Simonxag, via Wikimedia Commons.
Creationists see the origin of the Giardia baramin as part of God’s very good creation thousands of years ago. Although we cannot be 100% sure, we see not only non-pathogenic subspecies of Giardia, but also examples of their being a beneficial part of an animal—and possibly human—microbiome. These scientific observations give credence to Giardia being a once-functional part of a GI ecosystem. In general, Giardia in a post-fallen world can be attributed to its modification and degeneration of its original form, displacement from original animals into humans, and its overwhelming numbers in the small intestine (Tables 3 and 4). In addition, a change in the ability of the immune system to handle the interaction of Giardia appears to influence disease state. Creation scientists maintain that a number of infectious/parasitic diseases can be explained by corruption of the original kind and sometimes may be due to a loss of genetic information that alters its original purpose.
| Commensal/Mutual | Pathogen |
|---|---|
| Minimal harm in some wildlife | Disease in dogs, cats |
| Minimal issues in 50% of children that are colonized early in life (discourages Clostridium [CDIF]) | Severe diarrhea in some children especially those with IgA deficiency |
| Immune stimulant | Immune deficient |
| Possible pre-Fall benefit | Post-Fall disease |
|
|
Table 4. “Symbiotic” Fast Facts
- Creation scientists suggest the original created purpose of Giardia was to both start and aid in the digestive processes of other gut microflora (microbiome).
- Past investigations have shown other flagellates, such as trypanosomes, can actually be beneficial to animals.
- Research has shown Giardia actually changes the normal microbiota of its host, promoting concentration of Proteobacteria (e.g. Escherichia coli) via enzymatic activity.
- Flagellated communalistic protozoa are associated with the microbiome of creatures as diverse as arthropods and primates.
- There are four species of bacteria in the gut in Australian termites critical for their survival and for digesting wood, so trypanosomes and Giardia also seem to be important for microbial health in some animals.
Locally Abundant
My students are finding Giardia locally in the James River (Lynchburg, VA), a pond near Liberty University science hall, Forest (VA), and Rustburg (VA). These protozoa are much easier to find using Giardia SNAP tests (Fig. 13) than by using traditional methods. We have found them locally abundant, and their occurrence correlates with local clinical cases. A local physician reports about 30–40 cases per year in the summer due to recreation on the James River in Virginia. We have local evidence of mallard ducks carrying Giardia, but see no evidence of disease in them. Ducks appear to be carrier of cysts, but do not transmit disease to humans.

Figure 13. Giardia SNAP photo. Positive results from Giardia rapid antigen test as indicated by line under the “T.” “C” line is the control. Image by Alan Gillen.
The probable pathogenesis for giardiasis (Bauman 2014) is its trophozoite masses that interfere with normal small intestinal absorption of chyme. Large quantities of undigested fatty foods are moved into the bowel and metabolized by bacteria leading to fowl-smelling hydrogen sulfide diarrhea (steatorrhea). This unfortunate condition is sometimes described as “explosive” due to methane/carbon dioxide pressure caused by this overabundant and atypical bacterial metabolism. Steatorrhea is the result of fat malabsorption. It can be chronic, especially in animals. Beavers are common carriers, but their symptoms are less extreme. Hence, when Giardia is passed to humans, it is called Beaver Fever. In dogs and cats, the diarrhea can be chronic and difficult.
Giardiasis is commonly transmitted by drinking water cross-contaminated with sewage. It is also known to exist in clear-running streams due to wildlife depositing their wastes in the water. To drink water from trout streams without boiling it or adding iodine tablets is unsafe. In other words, all an unlucky camper needs to do is swallow just one microscopic cyst and they are infected. Many unsuspecting campers and fishermen have picked up giardiasis when returning home from an outing. Instead of enjoying fresh, home-cooked fish, they instead experience diarrhea, gas, and abdominal pain. The moral of the story is to not drink untested waters from the wild.
The World Health Organization (WHO) reports an annual Giardia infection rate of approximately 200 million people in both developed and developing countries. Giardiasis is recognized as one of the top 10 most common parasitic diseases in the world. In the USA and Canada, 81% of raw surface waters contain Giardia. The study was done in 66 treatment plants in 14 states and in a province of Canada. Giardia was detected in 17% of filtered water from samples of these same treatment plants (Goater, Goater, and Esch 2014). It is usually the primary parasitic disease in the USA. Fortunately, it is seldom serious.
Summary and Conclusions
Giardia has two “faces” even in today’s world: a harmless commensal in some wildlife and a pathogenic parasite in humans and domestic animals. Giardia is a protozoan parasite of humans and animals that causes significant morbidity and diarrheal disease globally. Giardiasis is a zoonotic disease with widespread reservoirs in vertebrates. Parasites infect and complete their life cycle in infected wildlife, then shed Giardia cysts into water supplies. Over one billion people are estimated to have acute or chronic giardiasis, and rates of giardiasis approach 90% in some developing nations. When common, giardiasis has been implicated as a primary cause of retarding development for children, resulting in long-term consequences such as stunted growth. In the USA, it is the number one parasitic disease most years.
Despite its pathogenic effects in some, Giardia acts an ecosystem engineer (Barash et. al. 2017), indirectly modulating the availability of resources to other species in the same environment, and excreting novel waste products such as lipids or amino acids that can be metabolized by good, enteric bacteria. An ecological perspective on giardiasis could inform our understanding of Giardia’s pathogenesis. Some “very good” microbes in the mammalian gut benefit the host by breaking down complex substrates, supplying essential nutrients, and defending against opportunistic pathogens. Commensal microbes from wildlife account for about 109 cells/g of total microbes in the small intestine and 1012 cells/g in the large intestine (Barash et. al. 2017). The diversity of bacteria is critical for overall stability of the gut. This microbial diversity can confer resilience and flexibility of commensal microbial responses to various external stresses, promoting the interface of the immune system (Gillen and Conrad 2014). Commensal gut microbial communities are known to both limit and control pathogen colonization, and thus the disruption of the gut ecosystem may impact consequent symptoms of diarrheal disease.
The creation model states that God created during the six-day creation week. After that there were no more creative acts. Therefore some creation biologists feel parasites, like Giardia, must be a modified form of life that was originally complete and that assisted with good symbiotic associations in the gut microbiome (along with E. coli) prior to the Fall. When God cursed the earth (Genesis 3), some protozoan animals eventually lost their original beneficial function due to loss of genetic information, displacement from its original home, and they developed dependence on other creatures—in other words, became parasitic with associated pathogenesis. Although the original design may be effaced, the intricate Giardia “cute” face and its interwoven complexity has not been erased. It is a wondrous, wild creation.
References
Barash NR, Maloney JG, Singer SM, and Dawson SC. 2017. Giardia Alters Commensal Microbial Diversity Throughout the Murine Gut. Infect. Immun. doi:10.1128/IAI.00948-16.
Bauman R. 2014. Microbiology with Diseases by Body System, 3rd ed. San Francisco, CA: Pearson Benjamin/Cummings Pub. Co.
Behe MJ. 2006. Darwin’s Black Box: The Biochemical Challenge to Evolution. New York, NY: Free Press.
Dobell C. 1932. Anthony van Leeuwenhoek and His “Little Animals.” London: Staples Press.
Gillen AL. 2014. The Genesis of Germs: Disease and the Coming Plagues in a Fallen World. Green Forest, AK: Master Books.
Gillen AL and Oliver D. 2012. “Antony van Leeuwenhoek: Creation ‘Magnified’ through His Magnificent Microscopes.” Answers In-Depth. (Aug. 15, 2012) https://answersingenesis.org/creation-scientists/profiles/antony-van-leeuwenhoeks-microscopes-creation-magnified/.
Gillen, AL and Conrad J. 2014. “Our Impressive Immune System: More than a Defense.” (Posted January, 15, 2014) Answers In-Depth. https://answersingenesis.org/human-body/our-impressive-immune-system-more-than-a-defense/.
Goater T, Goater C, and Esch G. 2014. Parasitism: The Diversity and Ecology of Animal Parasites. 2nd Edition. Cambridge, UK: Cambridge University Press.
Lenaghan SC, Davis CA, Henson WR, Zhang Z, and Zhang M. 2011. {High-speed Microscopic Imaging of Flagella Motility and Swimming in Giardia lamblia trophozoites. Proceedings of the National Academy of Sciences of the United States of America, 108(34), 13891–13892. Retrieved from doi:10.1073/pnas.1106904108.
Lincicome DR, 1971. “The Goodness of Parasitism: a New Hypothesis.” In Cheng TC. Some Aspects of the Biology of Symbiosis. Baltimore: University Park Press. Pages 139–227.
Lincicome DR and Sheppersox JR. 1963. “Increased Rate of Growth of Mice Infected with Trypanosoma duttoni.” Journal of Parasitology, 49, 31–34.
Marquardt WC, Demaree RS, and Grieve RB. 2000. Parasitology and Vector Biology, 2nd edition. San Diego, CA: Harcourt Academic Press.
Purdom G. (2012). “Magnificent Motors: God Invented It First.” Answers in Genesis. https://answersingenesis.org/biology/microbiology/magnificent-motors/.
Roberts LS, Janovy J Jr., and Nadler S. 2013. Schmidt and Roberts’ Foundations of Parasitology, 9th edition. Boston, MA: WCB McGraw-Hill.



{kind=link}
{kind=link}
{kind=link}
._Natuurkundige_te_Delft_Rijksmuseum_SK-A-957.jpeg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Support the creation/gospel message by donating or getting involved!

Answers in Genesis is an apologetics ministry, dedicated to helping Christians defend their faith and proclaim the good news of Jesus Christ.
- Customer Service 800.778.3390
- © 2024 Answers in Genesis