Part 1 Introduction
Terminology
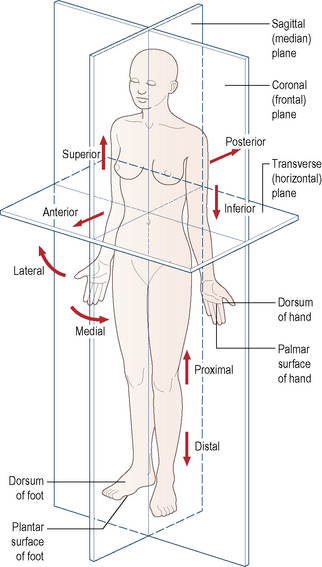
It is essential for students beginning their study of anatomy to become familiar with an internationally accepted vocabulary, allowing communication and understanding between all members of the medical and paramedical professions throughout the world. Perhaps the single, most important descriptive feature of this vocabulary is the adoption of an unequivocal position of the human body. This is known as the anatomical position. It is described as follows: the body is standing erect and facing forwards; the legs are together with the feet parallel so that the toes point forwards; the arms hang loosely by the sides with the palm of the hand facing forwards so that the thumb is lateral (Fig. 1.1). All positional terminology is used with reference to this position, irrespective of the actual position of the body when performing an activity.
The following is a list of more commonly used terms which describe the position of anatomical structures:
To facilitate the understanding of the relation of structures one to another and the movement of one segment with respect to another, imaginary reference planes pass through the body in such a way that they are mutually perpendicular to each other (Fig. 1.1). Passing through the body from front to back and dividing it into two symmetrical right and left halves is the sagittal (median) plane. Any plane parallel to this is also known as a parasagittal (paramedian) plane.
Within each plane a single axis can be identified, usually in association with a particular joint, about which movement takes place. An anteroposteriorly directed axis in the sagittal (or a paramedian) plane allows movement in a coronal plane. Similarly, a vertical axis in a coronal plane allows movement in a transverse plane. Lastly a transverse (right to left) axis in a coronal plane provides movement in a paramedian plane.
By arranging that these various axes intersect at the centre of joints, the movements possible at the joint can be broken down into simple components. It also becomes easier to understand how specific muscle groups produce particular movements, as well as to determine the resultant movement of combined muscle actions.
Terms used in describing movement
Rarely do movements of one body segment with respect to another take place in a single plane. They almost invariably occur in two or three planes simultaneously, producing a complex pattern of movement. However, it is convenient to consider movements about each of the three defined axes separately. Movement about a transverse axis occurring in the paramedian plane is referred to as flexion and extension; that about an anteroposterior axis in a coronal plane is termed abduction and adduction; and finally, that about a vertical axis in a transverse plane is termed medial and lateral rotation.
All movements are described, unless otherwise stated, with respect to the anatomical position, this being the position of reference. In this position joints are often referred to as being in a ‘neutral position’.
Flexion and extension of the foot at the ankle joint may be referred to as plantarflexion and dorsiflexion respectively.
Supination and pronation are terms used in conjunction with the movements of the forearm and foot.
Inversion and eversion are terms used to describe composite movements of the foot.
Nervous system
Introduction
The nervous system consists of highly specialized cells designed to transmit information rapidly between various parts of the body. Topographically it can be divided into two major parts: the central nervous system (CNS) and the peripheral nervous system (PNS). The brain and spinal cord constitute the CNS, which lies within the skull and vertebral canal (see p. 572 and p. 489), while nerves in the PNS connect the CNS with all other parts of the body (see p. 7).
The PNS consists of cells that connect the CNS with the other tissues of the body. These cells are aggregated into a large number of cable-like structures called nerves, which are threaded like wires throughout the tissues of the body.
Cellular structure
The basic cellular unit of the nervous system is the nerve cell or neuron, which differ in size and shape according to their function and location within the nervous system. All neurons have three characteristic components: a cell body, an axon, and dendrites (Fig. 1.2A).
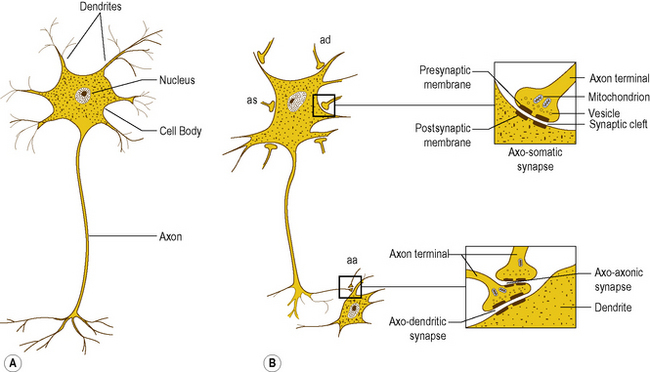
Figure 1.2 (A) Microscopic appearance of a neuron and its component parts, (B) neuron with multiple axo-somatic (as), axo-dendritic (ad) and axo-axonic (aa) synapses. The insets show the detailed structure of the synapse.
The cell body is an expanded part of the cell containing the nucleus and apparatus necessary to sustain the metabolic activities of the cell. The axon is a longitudinal, tubular extension (process) of the cell membrane and cytoplasm which transmits information away from the cell body. The cell membrane surrounding the axon is referred to as the axolemma. Dendrites are extensions of the cell membrane that radiate from the cell body in various directions, and are responsible for receiving information and transmitting it to the cell body.
Structurally, dendrites differ from axons as they typically undergo extensive branching close to the cell body, whereas axons remain singular for most of their course only branching at their terminal ends. A neuron has only one axon, but may have several dendrites; the lengths and calibre of axons and dendrites vary, depending on the particular function of the neuron.
Interneural connections
Individual neurons convey information by conducting electrical action potentials along their cell membrane, with communication between separate neurons occurring chemically at a specialized structure called a synapse.
Synapses are formed by the close approximation of a small discrete area of the cell membrane of one neuron to a reciprocal area of the membrane of a second neuron (Fig. 1.2B). The apposed membranes are specially modified and are separated by a narrow gap about 0.02 μm in width, called the synaptic cleft. Across the synapse one cell communicates with the next, with communication being unidirectional. The membrane of the cell transmitting the information is the presynaptic membrane, and that of the cell receiving the information the postsynaptic membrane.
Synapses typically occur between the axon of one neuron and the dendrite of another. However, they may also occur between axons and cell bodies, axons and axons, and even between dendrites and dendrites. Some neurons receive only a few synapses while others may receive thousands.
The purpose of synapses is not to relieve one neuron of information and simply pass it on to the next, but to allow the interaction of information from perhaps several diverse sources on a single neuron. The activity of a given neuron may be influenced by many others and, conversely, by having several terminal branches to its axon, a single neuron may influence many other neurons.
By being connected to one another in diverse ways, groups of neurons are organized to subserve different functions of the nervous system. The patterns of connections, or circuits, vary in complexity and, in general, the more sophisticated the function being subserved, the more complex the circuitry.
Myelination
Myelination is a process whereby individual axons are wrapped in a sheath of lipid called myelin. The myelin sheath serves as a protective and insulating coating for the axon, and enhances the speed of conduction of electrical impulses along it.
In the PNS, myelination is by cells called Schwann cells: it is achieved by a Schwann cell curling an extension of its cell membrane around the shaft of the axon in a spiral manner (Fig. 1.3A). As the Schwann cell extension wraps around the axon, the cytoplasm in it is squeezed out until only a double layer of the cell membrane of the Schwann cell remains spiralling around the axon (Fig. 1.3A). In this way the myelin sheath is formed by the lipid and protein of the cell membrane of the Schwann cell that surrounds the axon. Peripheral to the myelin sheath, the axon is surrounded by the cytoplasm of the Schwann cell, with the outermost cell membrane of the Schwann cell acting as a second membrane to the axon; this forms the neurolemma (Fig. 1.3A).
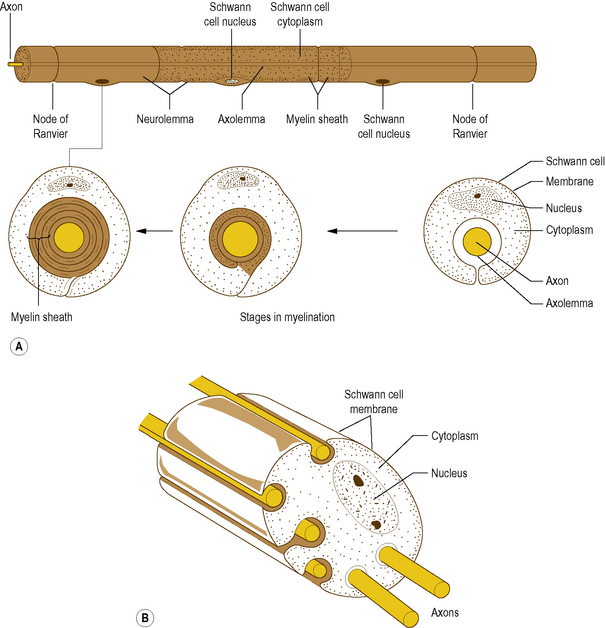
Figure 1.3 (A) Myelinated axon in the peripheral nervous system, (B) relationship between unmyelinated axons and Schwann cells.
Along its length, a given axon is ensheathed in series by a large number of Schwann cells, but each Schwann cell is related only to a single axon (Fig. 1.3A). Junction points where the myelin sheath of one Schwann cell ends and the sheath of the next begins are known as Nodes of Ranvier. An axon together with the myelin sheath and the Schwann cells that surround it is referred to as a nerve fibre.
While many axons in the PNS are myelinated a large number remain unmyelinated. Instead of being enveloped by a tightly spiralling sheath of myelin, they run embedded in invaginations of Schwann cell membranes, so that a single Schwann cell may envelop several axons (Fig. 1.3B). Unmyelinated axons are afforded less physical protection than myelinated ones, but the foremost difference between them is that unmyelinated axons conduct impulses at much slower velocities.
In the CNS, myelination is performed by specialized cells called oligodendrocytes. The process is basically similar to that in the PNS, except that a given oligodendrocyte is usually involved in the simultaneous myelination of several separate axons (Fig. 1.4). Unmyelinated axons in the CNS run embedded in cytoplasmic extensions, or processes, of oligodendrocytes.
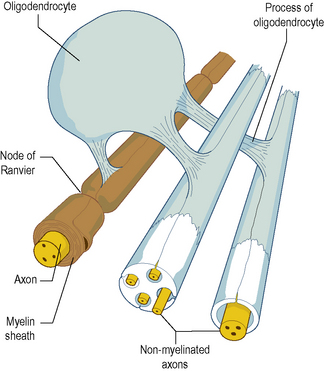
Figure 1.4 Relationship between an oligodendrocyte and the several myelinated and unmyelinated axons it ensheathes.
Several diseases may affect myelinating cells, including toxic and metabolic diseases, and most notably multiple sclerosis. In these diseases, neurons are not necessarily directly affected, but the loss of their myelin covering affects the way they conduct action potentials resulting in disordered neural function.
Structure of a peripheral nerve
A peripheral nerve is formed by the parallel aggregation of a variable number of myelinated and unmyelinated axons: the greater the number of axons, the larger the size of the nerve. Microscopically, within a peripheral nerve, myelinated axons are surrounded by their individual Schwann cell sheaths and unmyelinated axons run embedded in invaginations of the Schwann cell membrane. The axons are held together by sheaths of fibrous tissue which constitute additional coatings that protect them from external mechanical and chemical insults.
Individual myelinated axons are surrounded by a tubular sheath of fibrous tissue, the endoneurium, whereas clusters of axons are held together by a larger fibrous sheath, the perineurium (Fig. 1.5). Unmyelinated axons are not enclosed by endoneurium but run in isolated bundles parallel to myelinated axons and are enclosed with them in the perineurial sheath.
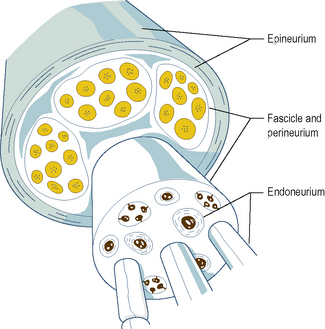
A bundle of axons enclosed within a single perineurial sheath is referred to as a nerve fascicle (Fig. 1.5). Axons within a fascicle largely remain within that same fascicle throughout the course of the nerve until their peripheral distribution. However, along the course of a peripheral nerve axons may at times leave one fascicle to enter and continue within an adjacent fascicle.
The fascicles within a peripheral nerve are bound together by an external sheath of fibrous tissue, the epineurium (Fig. 1.5), which forms the external surface of the macroscopic nerve. As a peripheral nerve passes through the tissues of the body, it gives off branches composed of one or more fascicles of the parent nerve, which leave it to reach their particular destination. Along the course of a nerve, this process is repeated until all the fascicles and axons in the nerve have been distributed to their target tissues.
The peripheral nervous system
Nerves that supply the structural tissues, such as bone, muscle and skin, are referred to as somatic nerves. Groups of somatic nerves innervate specific areas or regions and are covered in detail in the respective region.
The nerves that supply the viscera, such as the heart, lungs and digestive tract, are referred to as visceral nerves. However, because the nervous functions concerning viscera are largely automatic and subconscious, that part of the nervous system that innervates viscera is referred to as the autonomic nervous system (ANS). It has components in both the central and PNSs: it is described in detail on page 500.
Constituents of peripheral nerves
Peripheral nerves are composed of different types of axons that are classified according to their size, function or physiological characteristics. The broadest classification of axons recognizes afferent (or sensory) fibres and efferent (or motor) fibres. The terms ‘afferent’ and ‘efferent’ refer to the direction in which axons conduct information: afferent fibres conduct towards, while efferent fibres conduct away from the CNS. The term ‘sensory’ refers to axons that convey information to the CNS about events that occur in the periphery; ‘motor’ fibres cause events in the periphery, usually in the form of contraction of voluntary or smooth muscle.
Axons may also be classified according to their conduction velocities, which in turn are proportional to their sizes (Fig. 1.6). If a peripheral nerve is stimulated electrically and recordings of the evoked activity are taken some distance along the nerve, a wave of electrical activity can be recorded. The wave is generated by the summation of the electrical activity in each of the axons in the nerve, its size being proportional to the number of axons in the nerve, while its shape reflects the type of axons present.
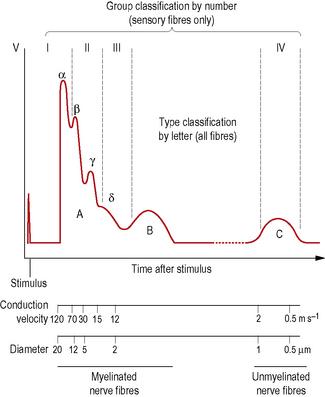
Figure 1.6 Neurogram of an idealized peripheral nerve. Activity in different types of nerve fibres is reflected in depolarizations that occur at different times after the delivery of an electrical stimulus to the nerve.
The waveform obtained from a nerve containing every known type of axon is represented in Fig. 1.6. It has three principal peaks, named the A, B and C waves. The A wave occurs very soon after the triggering stimulus, as it is produced by rapidly conducting axons with velocities in the range 12–120 ms−l: these are classified as A fibres. The A wave is broken down into several secondary peaks called Aα, Aβ, Aγ and Aδ waves, each being produced by a particular subgroup of rapidly conducting axons.
Not all peripheral nerves contain every type of fibre, therefore not all nerves will exhibit A, B and C waves. For example, B fibres are not regularly present in all nerves; B waves are therefore not always recordable.
Myelinated axons conduct impulses faster than unmyelinated ones, and among myelinated axons those with larger diameters conduct impulses faster than those with smaller diameters. The conduction velocity of a myelinated axon is directly proportional to the total diameter of the axon and its myelin sheath. A and B fibres are myelinated axons of various diameters and fast conduction velocities, while C fibres are unmyelinated axons with small diameters and slow conduction velocities.
Functionally, Aα and Aγ fibres represent motor fibres connected to voluntary muscles and also include certain sensory fibres that transmit position sensation from skeletal muscles. Aβ fibres mediate the sensations of touch, vibration and pressure from skin; Aδ fibres are sensory fibres that mediate pressure, pain and temperature sensations from skin, pain and pressure from muscle, and pain, pressure and position sensation from ligaments and joints. B fibres are preganglionic sympathetic efferent fibres. C fibres are largely unmyelinated sensory fibres that arise in virtually all tissues of the body and transmit pain, temperature and pressure sensations, however, some are postganglionic sympathetic neurons. Because Aβ fibres have large diameters they are sometimes referred to as large diameter afferent fibres, while Aδ and C fibres are referred to collectively as small diameter afferent fibres.
Another system of axon classification relates specifically to sensory fibres. In this system, sensory fibres are classified according to their conduction velocities into groups I, II, III and IV. These groups have the same conduction velocities as type Aα, Aβ, Aδ and C fibres respectively, as shown in Fig. 1.6, but do not include the Aα and Aβ motor fibres. The fibres of groups III and IV largely mediate pain and temperature sensations. Group II constitutes fibres that mediate pressure and touch, and fibres that form spray endings in muscle spindles (p. 15). Group I is divided into groups Ia and Ib. Group Ia fibres are slightly larger and innervate muscle spindles, while group Ib fibres innervate Golgi tendon organs (p. 15).
Nerve endings
The terminals of axons in peripheral nerves have unique structures depending on their function. Motor axons have terminals designed to deliver a stimulus to muscle cells (see p. 16), while sensory axons have terminals designed to detect particular types of stimuli. These terminals are called receptors, and a diversity of morphological types can be found (see p. 26).
Nervous system
Components of the musculoskeletal system
As this book is concerned essentially with the musculoskeletal system a brief account of the major tissues of the system, i.e. connective, skeletal and muscular tissue, and of the type of joints which enable varying degrees of movement to occur, is given as it will aid in understanding the mobility and inherent stability of various segments. The initiation and coordination of movement is the responsibility of the nervous system.
Connective tissue
Connective tissue is of mesodermal origin and in the adult has many forms; the character of the tissue depends on the organization of its constituent cells and fibres.
Fat
Fat is a packing and insulating material; however, in some circumstances it can act as a shock absorber, an important function as far as the musculoskeletal system is concerned. Under the heel, in the buttock and palm of the hand, the fat is divided into lobules by fibrous tissue septa, thereby stiffening it for the demands placed upon it.
Fibrous tissue
Fibrous tissue is of two types. In white fibrous tissue there is an abundance of collagen bundles, whereas in yellow fibrous tissue there is a preponderance of elastic fibres.
White fibrous tissue is dense, providing considerable strength without being rigid or elastic. It forms: (1) ligaments, which pass from one bone to another in the region of joints, uniting the bones and limiting joint movement; (2) tendons for attaching muscles to bones; and (3) protective membranes around muscle (perimysium), bone (periosteum) and many other structures.
Yellow fibrous tissue, on the other hand, is highly specialized, being capable of considerable deformation yet returning to its original shape. It is found in the ligamenta flava associated with the vertebral column as well as in the walls of arteries.
Skeletal tissue
Skeletal tissues are modified connective tissues, in which the cells and fibres have a particular organization and become condensed so that the tissue is rigid.
Cartilage
Cartilage is supplementary to bone, forming wherever strength, rigidity and some elasticity are required. In fetal development, cartilage is often a temporary tissue, being later replaced by bone. However, in many places cartilage persists throughout life. Although a rigid tissue, it is not as hard or strong as bone. It is also relatively non-vascular, being nourished by tissue fluids. A vascular invasion of cartilage often results in the death of the cells during the process of ossification of the cartilage and its eventual replacement by bone. Except for the articular cartilage of synovial joints, cartilage possesses a fibrous covering layer, the perichondrium.
There are three main types of cartilage: hyaline cartilage, white fibrocartilage and yellow fibrocartilage.
Hyaline cartilage
Hyaline cartilage forms the temporary skeleton of the fetus from which many bones develop. Its remnants can be seen as the articular cartilages of synovial joints, the epiphyseal growth plates between parts of an ossifying bone during growth, and the costal cartilages of the ribs. At joint surfaces it provides a limited degree of elasticity to offset and absorb shocks, as well as providing a relatively smooth surface permitting free movement to occur. With increasing age, hyaline cartilage tends to become calcified and sometimes ossified.
White fibrocartilage
White fibrocartilage contains bundles of white fibrous tissue which give it great tensile strength combined with some elasticity so that it is able to resist considerable pressure. It is found at many sites within the musculoskeletal system: (1) within the intervertebral discs between adjacent vertebrae; (2) in the menisci of the knee joint; (3) in the labrum surrounding and deepening the glenoid fossa of the shoulder joint and the acetabulum of the hip joint; (4) in the articular discs of the radiocarpal (wrist), sternoclavicular, acromioclavicular and temporomandibular joints; and (5) as the articular covering of bones which ossify in membrane, e.g. the clavicle and mandible.
Bone
Bone is extremely hard with a certain amount of resilience. It is essentially an organic matrix of fibrous connective tissue impregnated with mineral salts. The connective tissue gives the bone its toughness and elasticity, while the mineral salts provide hardness and rigidity, the two being skilfully blended together. The mineral component provides a ready store of calcium, which is continuously exchanged with that in body fluids, with the rate of exchange and overall balance of these mineral ions being influenced by several factors including hormones.
Each bone is enclosed in a dense layer of fibrous tissue, the periosteum, with its form and structure adapted to the functions of support and resistance of mechanical stresses. Being a living tissue, bone is continually remodelled to meet these demands; this is particularly so during growth. The structure of any bone cannot be satisfactorily considered in isolation; it is dependent upon its relationship to adjacent bones and the type of articulation between them, as well as the attachment of muscles, tendons and ligaments to it.
The internal architecture of bone reveals a system of struts and plates (trabeculae) running in many directions (Fig. 1.7), which are organized to resist compressive, tensile and shearing stresses. Surrounding these trabecular systems, which tend to be found at the ends of long bones, is a thin layer of condensed or compact bone (Fig. 1.7). The network of the trabeculae, because of its appearance, is known as cancellous or spongy bone. In the shaft of a long bone there is an outer, relatively thick ring of compact bone surrounding a cavity, which in life contains bone marrow.
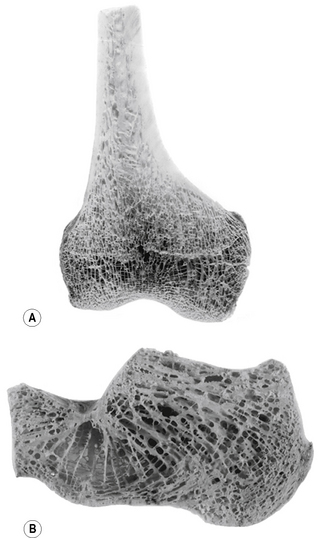
Figure 1.7 Trabecular arrangement within bone: (A) coronal section through the lower end of the femur, (B) sagittal section through the calcaneus.
Red and white blood cells are formed in red bone marrow, which after birth is the only source of red blood cells and the main source of white blood cells. In infants, the cavities of all bones contain red marrow. However, this gradually becomes replaced by yellow fat marrow, so that at puberty red marrow is only found in cavities associated with cancellous bone. With increasing age many of these red marrow containing regions are replaced by yellow marrow. Nevertheless, red marrow tends to persist throughout life in the vertebrae, ribs and sternum, and the proximal ends of the femur and humerus.
For descriptive purposes bones can be classified according to their shape:
Both irregular and short bones consist of a thin layer of compact bone surrounding cancellous bone.
Bone development
Bone develops either directly in the mesoderm by the deposition of mineral salts, or in a previously formed cartilage model. When the process of calcification and then ossification takes place without an intervening cartilage model, it is known as intramembranous ossification, with the bone being referred to as membrane bone. However, if there is an intervening cartilage model, the process is known as endochondral ossification, with the bone being referred to as cartilage bone. This latter process is by far the most common.

Full access? Get Clinical Tree


