
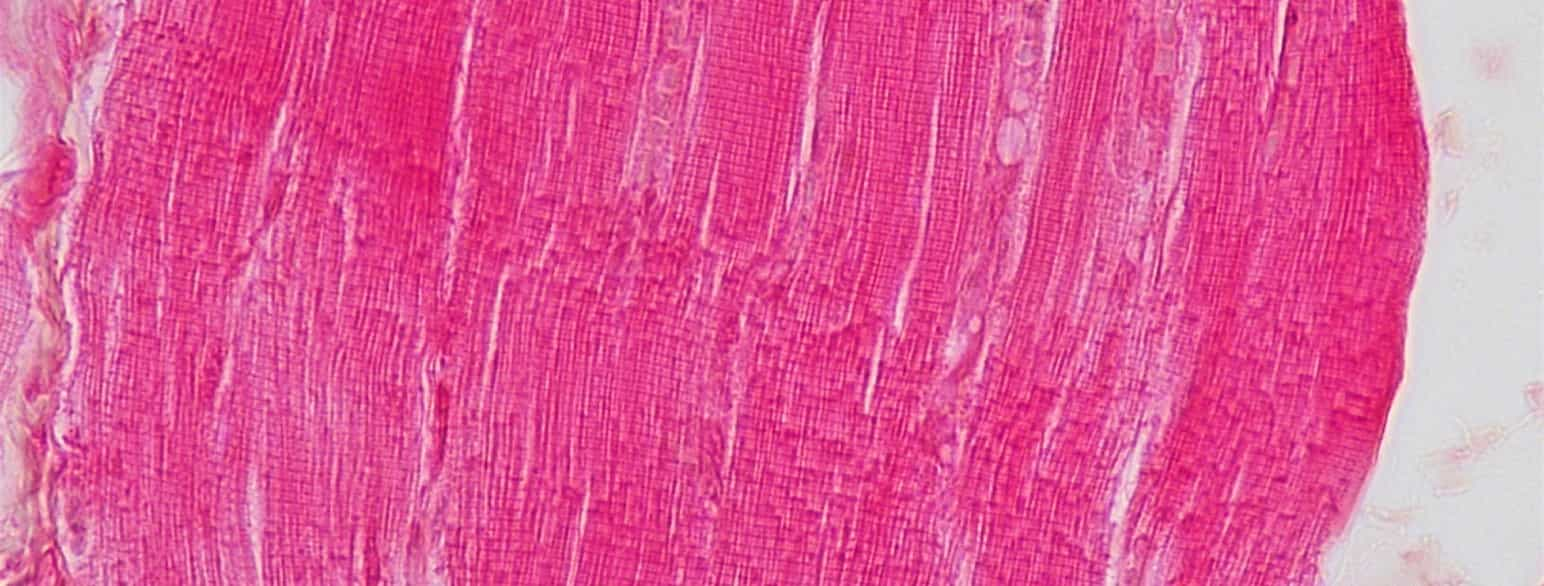
Muskelvev består av celler som er spesialiserte til å trekke seg sammen (kontrahere). Fordi cellene er langstrakte, kalles de også muskelfibrer. På grunnlag av forskjeller i cellenes utseende, egenskaper og funksjon deles muskelvev inn i tre grupper:
- tverrstripet skjelettmuskulatur
- glatt muskulatur
- hjertemuskulatur
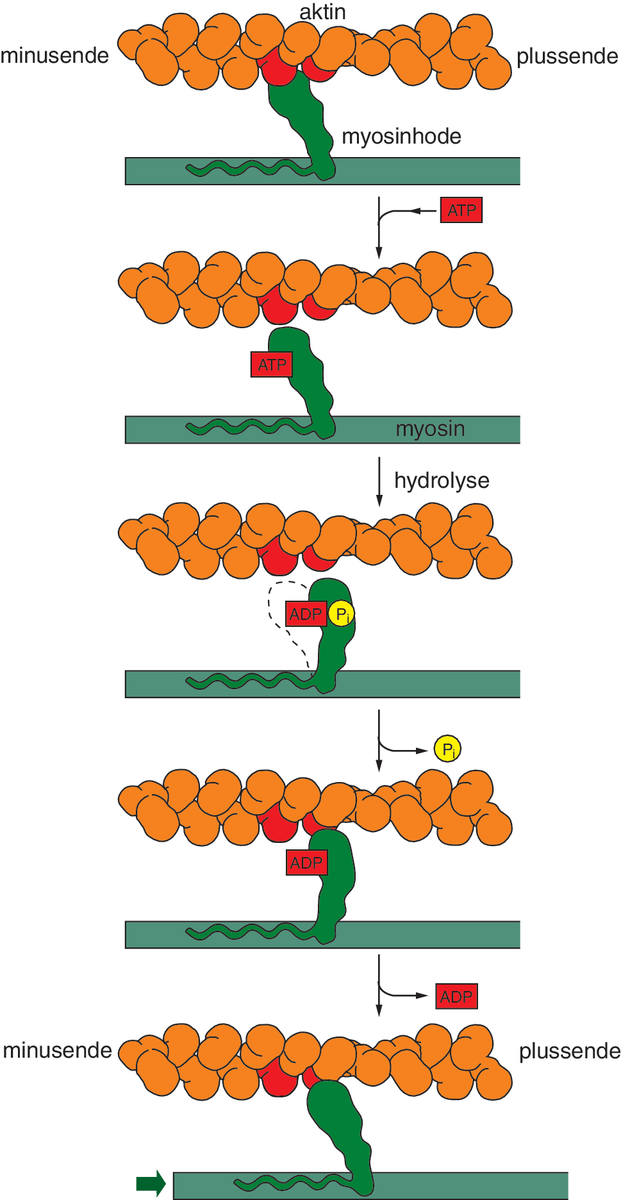
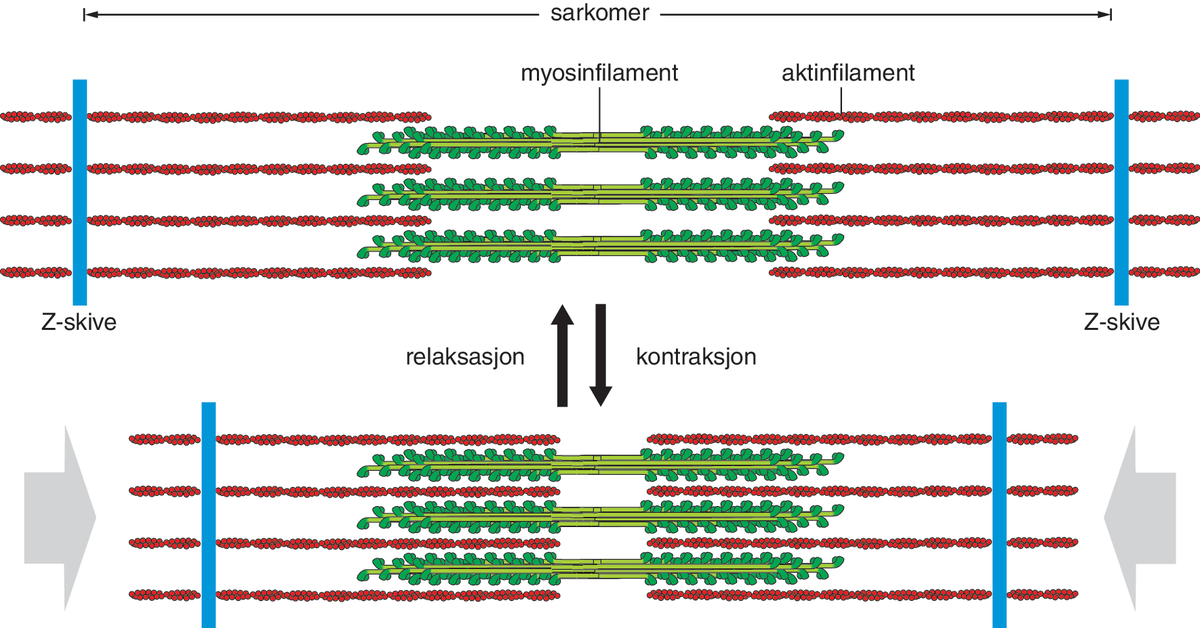
I alle tre skyldes kontraksjonen glidebevegelser av filamenter (tynne, trådformede strukturer) i forhold til hverandre. Det er to hovedtyper filamenter, aktin- og myosinfilamenter. Aktinfilamentene er bygd opp av det globulære (kuleformede) proteinet aktin, som henger etter hverandre som perler på en snor. I hver tråd ligger alle aktinmolekylene vendt samme vei, slik at tråden får retning, kalt pluss- og minusretning, og endene av tråden er henholdsvis kalt pluss- og minusende.
Myosinfilamentene består av myosin-II, et langstrakt protein med globulært hode og lang hale. Mange myosin-II-molekyler ligger pakket sammen i bunter, litt forskutt i forhold til hverandre og med hodene pekende ut mot omgivende aktintråder. Myosintrådene får dermed utseende som en flaskebørste, men med to «børster» pekende i motsatt retning, hektet sammen i halen. Myosin er et motorprotein, som kan omdanne kjemisk energi til mekanisk arbeid. Ved kontraksjon vandrer myosin i plussretning langsetter aktinfilamentene, mens det spalter adenosintrifosfat (ATP), som utgjør energikilden. Fordi myosinmolekylene i buntene henger fast i halene til hverandre og peker i motsatt retning, blir effekten at aktintrådene trekkes mot midten av myosinbunten.
I tverrstripede muskelceller styres kontraksjonen gjennom regulering av myosinhodenes tilgang til aktin. I avslappet tilstand blokkerer lange trådformede proteiner bindingen mellom myosin og aktin. Nervesignaler utløser elektriske signaler i muskelcellemembranen. Dette forårsaker kortvarig frigjøring av kalsiumioner inne i muskelcellen. Disse binder seg til regulerende proteiner som trekker de blokkerende trådene vekk slik at myosin kan bindes til aktin.

Kommentarer
Kommentarer til artikkelen blir synlig for alle. Ikke skriv inn sensitive opplysninger, for eksempel helseopplysninger. Fagansvarlig eller redaktør svarer når de kan. Det kan ta tid før du får svar.
Du må være logget inn for å kommentere.