
Preprint
Review
Why Hydrogen Peroxide-Producing Proteins are not suitable Targets for Drug Development
Altmetrics
Downloads
230
Views
38
Comments
0



This version is not peer-reviewed
Submitted:
26 March 2024
Posted:
27 March 2024
You are already at the latest version
Alerts
Abstract
Reactive oxygen species (ROS) play an important role in biological processes and offer hope for the development of effective treatments for ROS-dependent diseases, including cancer. The interaction of hydrogen peroxide with proteins stimulates the creation of chemical molecules that can weaken or stop the pathological process in cancer. However, reliable drugs have not yet been described. To understand the reason for the failure, we studied the behavior of ROS-containing compounds in cancer cells. The epidermal growth factor receptor (EGFR) is activated by interaction with a ligand protein, as well as by binding the chemical structure of hydrogen peroxide to the catalytic cysteine in the intracellular domain. Both mechanisms trigger signaling pathways in cells. The EGFR can be inactivated by protein tyrosinase phosphatase PTP-1B, which itself is activated by hydrogen peroxide. A simple in-gel fluorescence technique was used to demonstrate the rapid binding of hydrogen peroxide to a large number of proteins, including both activating and inactivating target proteins. Natural defense systems take longer to action and apparently cannot prevent hydrogen peroxide from binding to protect cells from exposure to unwanted proteins. Therefore, superoxide dismutase, which produces hydrogen peroxide, is not a suitable target for disease therapy.
Keywords:
Subject: Medicine and Pharmacology - Oncology and Oncogenics
Introduction
Global ionizing radiation and photochemical reactions generate free radicals, suggesting that the evolution of oxygen-consuming organisms was driven by their ability to tolerate oxygen free radicals. A striking example of such evolution is the birth of a child, which represents the transition of the organism from a hypoxic maternal environment to the oxygen environment necessary for life. The action of free radicals requires a better understanding of the molecular mechanisms of tolerance, especially in the face of growing threats of climate change and military excesses.
The high redox potential of oxygen, which has two unpaired electrons in its outer orbit, allows it to serve as an ideal electron acceptor. The reduction of dioxygen (O2) to superoxide radical (O2•-) converts oxygen into free radicals known as reactive oxygen species (ROS), which attack various targets in the human body (Figure 1) [1]. The enzyme superoxide dismutase (SOD) was discovered as an important protein in the first line of cell defense against oxygen free radicals [2]. Among the three classes of SODs that have a distinct subcellular localization in eukaryotes, Cu/Zn SODs are expressed in the cytoplasm and extracellularly, and MnSODs are expressed in mitochondria [3]. SOD directs the reaction of superoxide anion (O2•) with the formation of hydrogen peroxide (H2O2) and oxygen (O2) according to the following reaction (2O2•- + 2H+ → H2O2 + O2) [4]. The resulting superoxide radical (O2•-) is converted into a non-radical species with a high oxidation potential - hydrogen peroxide (H2O2), which can subsequently be transformed into a hydroxyl radical (HO•). Hydrogen peroxide decomposes into water and oxygen upon heating or in the presence of numerous substances, particularly salts of such metals as iron, copper, manganese, etc.
Three SOD isoforms are involved in many cellular functions, including the control of crosstalk in the cellular microenvironment, which has led to their extensive testing as possible therapeutic agents [5]. The presence of metal-containing cofactors in SOD isoforms allows the cell to maintain homeostasis and coordinate ROS signals between cellular compartments [6]. ROS generated in healthy cells can modify the functional building blocks of proteins and lipids, thereby preventing cell damage. However, in cancer cells, up- and down-regulation appears to be dysregulated, making ROS a hot topic in cancer research and anticancer drug development [7].
Both ligand-dependent and ligand-independent trans-activations of the transmembrane epidermal growth factor receptor (EGFR) have been described, indicating the high complexity of this signaling system. Non-radical H2O2 acts as a second messenger and plays a critical role in cellular functions, including signaling and metabolic pathways [8]. Oxidative stress theory suggests that ROS underlie disease states, which has prompted the proposal of in vitro and in vivo models to demonstrate their role in the initiation and progression of human diseases [9].
Reactive oxygen species play an important role in biological processes in human body and provide hope for the development of effective treatments for ROS-dependent diseases. However, given the virtual lack of progress in developing drugs to counteract the negative effects of ROS, the validity of considering them as reliable therapeutic agents targeting SOD-producing proteins has been questioned.
2. Results
2.1. The Role of EGFR in Redox Processes
In humans, 58 proteins belong to the family of receptor tyrosine kinases (RTKs), involved in the phosphorylation of transmembrane proteins in cellular processes [10]. All RTKs are activated by a unique mechanism consisting of homodimerization and heterodimerization of the extracellular domain followed by autophosphorylation of tyrosine residues located at the C-terminus of the intracellular domain [11]. Phospho-tyrosine residues can be recognized by proteins bearing so-called SH2 (SRC homology domain 2) and PTB (phospho-tyrosine binding domain) domains, which can implement the signal either by phosphorylation (e.g. JAK/STAT) or by recruiting docking proteins that initiate signaling cascades (e.g., RAS/MAPK and PI3K/AKT) in respective pathways as demonstrated for EGFR [12] (Figure 2). The enormous amount of research carried out on the EGFR receptor since its discovery by Stanley Cohen, who together with Rita Levi-Montalcini received the Nobel Prize in 1986, has led to an understanding of the role of this attractive protein in many life processes such as survival, signaling pathways, death [13].
Phosphorylation of proteins in the cell is regulated by the activity of protein kinase and phosphatase, which can be modulated by intracellular hydrogen peroxide through various mechanisms. The interaction of the cognate ligand EGF with the extracellular domain of EGFR promotes dimerization of the receptor, which is necessary for the activation of the ATP-binding catalytic site located in the cytoplasmic domain [14]. Hydrogen peroxide was found to enhance EGFR tyrosine phosphorylation in human squamous cell carcinoma cells, whereas stimulation with EGF ligand triggered tyrosine phosphorylation at the receptor [15]. In addition, inhibition of Cys-dependent protein tyrosine phosphatases (PTPs) by hydrogen peroxide was found to be a critical mechanism inducing EGFR tyrosine phosphorylation [16]. The intracellular kinase domain of EGFR contains six Cys residues, and Cys797 is located in close proximity to the ATP-binding pocket and is preferentially exposed to endogenous hydrogen peroxide [17]. Therefore, oxidation of catalytic Cys797 in EGFR is responsible for the subsequent activation of downstream signaling pathways in cells [18,19].
Phosphorylation of EGFR is inhibited by protein tyrosine phosphatases, among which PTP-1B is the main dephosphorylating enzyme [20]. Phosphorylation of the receptor is associated with inactivation of PTP-1B by H2O2 [21] and results from the oxidation of the nucleophilic cysteine to the unstable electrophilic sulfenic acid (Cys-SOH), which causes cyclization with the adjacent amide group, resulting in the formation of inactive 3-isothiazolidinone at the PTP-1B catalytic site [22]. Hydrogen peroxide produced when EGF binds to EGFR oxidizes the catalytic Cys797 at the ATP binding site to sulfenic acid, which enhances autophosphorylation of the receptor [23]. Unstable sulfenic acid is further oxidized to stable sulfinic acid (Cys-SO2H) without loss of receptor kinase activity [18,24].
In our study, to determine whether EGFR activation depends on the dimerization state of SOD1, receptor tyrosine phosphorylation was assessed using an siRNA interference experiment [25]. The amount of both monomeric and dimeric forms of SOD1 was significantly reduced in cells after 5 min of exposure to H2O2-producing compounds compared to non-transfected cells or cells treated with scrambled siRNA. Therefore, SOD1 activity is critical for EGFR-mediated tyrosine phosphorylation.
The binding of hydrogen peroxide to EGFR also activated subsequent signal transduction in the absence of EGF ligand in the growth medium, as observed by analyzing tyrosine phosphorylation in the downstream proteins pShc, pERK1/2, pGab1 and pCBL [25].
Two mechanisms may trigger EGFR signaling pathways that are important for cell function under different environmental conditions. EGFR is activated by binding to small protein ligands as well as exposure to the chemical structure of H2O2.
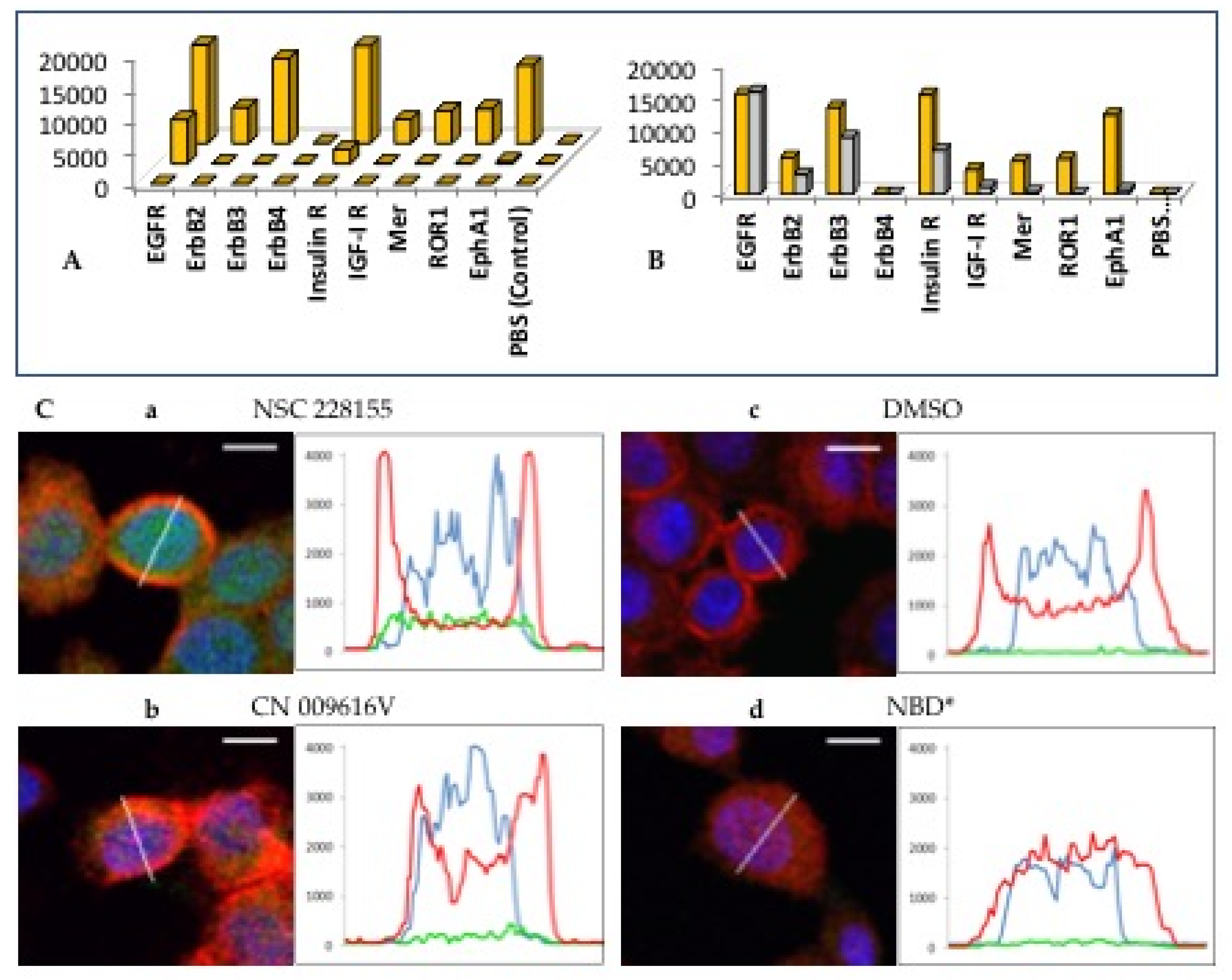
Fluorescent molecule is 4-nitro-2,1,3-benzoxadiazole (NBD), called nitrobenzofurazan, has an excitation wavelength (λex) of 440 nm and an emission wavelength (λem) of 480 nm and can interact with functional groups of proteins. Therefore, the NBD structure was used in the development of molecular probes [26] and fluorescent dyes for cell imaging [27,28]. The NBD scaffold carrying compound NSC 228155 induced phosphorylation of several RTKs, namely EGFR1, ErbB2, ErbB3, insulin R, IGF-IR, Mer, ROR1 and EphA1, within 10 minutes, as detected by microarray analysis of 40 proteins immobilized on a nitrocellulose membrane [25] (Figure 3A). In comparison, EGF ligand also showed a strong binding signal to EGFR, but weaker signal to other RTKs and no signal to ROR1 under the conditions used (Figure 3B). This suggests that both the EGF ligand and the NBD compound have a fairly similar ability to activate EGFR through different mechanisms, affecting the extracellular and intracellular domain of the receptor.
The sequential action of three enzymes, namely superoxide dismutase, catalase and glutathione peroxidase, usually protects cells from the action of toxic forms of ROS. Considering that the activity of these enzymes occurs in dimeric and possibly higher oligomeric forms, we assessed the oligomeric state of the corresponding enzymes in breast and prostate cancer cells treated with ROS [29]. It turned out that a significant part of the 16 kDa Cu/Zn-SOD1 monomer was combined into a 32 kDa dimeric structure in cells exposed to ROS for 5 min. At the same time, no changes in the molecular weight of catalase and glutathione peroxidase were detected in cells.
Lipophilic derivatives of NBD rapidly bind to SOD1, forming a stable dimeric protein in the absence of adequate catalase activity in cancer cells [29]. In the absence of a subsequent coupled redox reaction, non-reactive H2O2 should theoretically accumulate in the cytoplasm under the influence of SOD1. But H2O2 can be converted into a reactive form of ROS, namely the hydroxyl radical HO•, if the extracellular conditions change and catalase or another adequate oxidative reaction ensures this conversion.
The role of SOD1 in cell signaling was also proven for the DNA protein kinase catalytic subunit (DNA-PK) in prostate cancer cells exposed shortly to the compound NSC 228155 [30]. The DNA-PK is a key protein involved in the repair of DNA double-strand breaks, which are considered the most cytotoxic DNA lesions and result from endogenous events such as the production of ROS during cellular metabolism [31]. It was found that tyrosine phosphorylation as well as the expression and activity of DNA-PKcs drastically decreased in cells exposed to NBD compounds for 10 min. Of note is that this decrease was accompanied by increased protein ubiquitination and the activation of the proteasome machinery leading to the protein degradation [30].
Overproduction of ROS can lead to oxidative damage to a wide range of biomolecules, including nucleic acids, carbohydrates, lipids and proteins, which can trigger various cellular functions and the development of oxidative stress in the human body. The effects of electrophilic ROS on cellular signaling pathways typically occur through modulation of cytoplasmic kinase/phosphatase activity and alteration of nuclear transcription factor function in inflammatory diseases [32]. From a biological point of view, active electrophilic ROS can be considered as oxidative by-products formed during tightly controlled metabolic processes or during unregulated pathological processes. Substrate profiling experiments have shown that many enzymes can convert hundreds of substrates, highlighting the diversity of catalysts that utilize the active site in different metabolic transformations [33]. It is noteworthy to recall that metabolic enzymes are capable of accelerating chemical reactions by 26 orders of magnitude [34].
NBD compounds bind to EGFR and stimulate the formation of dimers, which is a necessary condition for further allosteric activation of the catalytic site in the cytoplasmic domain of the receptor. In this regard, NBD compounds behave as natural orthosteric agonists of EGFR, triggering signaling pathways similar to peptide modulators of other transmembrane receptors (for review, see [35]). Our data indicate that EGFR activation occurs through the binding of lipophilic NBD compounds to the intracellular domain of the receptor, resulting in H2O2 production by dimerized SOD1 in cancer cells. This binding promotes an excess of active SOD1 dimers without increasing the activity of subsequent reactions catalyzed by catalase and glutathione peroxidase. Once the concentration of hydrogen peroxide catalyzed by SOD1 reaches a critical level, the catalytic cysteine is sulfonylated in both EGFR and PTP-1B. Together, activated protein kinase and inactivated protein phosphatase mediate increased phosphorylation of EGFR. This suggests that SOD1 plays a key role in EGFR tyrosine phosphorylation, at least when cognate ligands are depleted in human cells. Hydrogen peroxide, through binding to the catalytic cysteine of EGFR, may act as a physiological mediator of signaling in cancer cells. These data suggest that both the EGF ligand and the NBD compound have a fairly similar ability to activate EGFR through different mechanisms, affecting the extracellular and intracellular domain of the receptor.
2.2. Straigthforward Detection of Hydrogen Peroxide Binding to Many Proteins
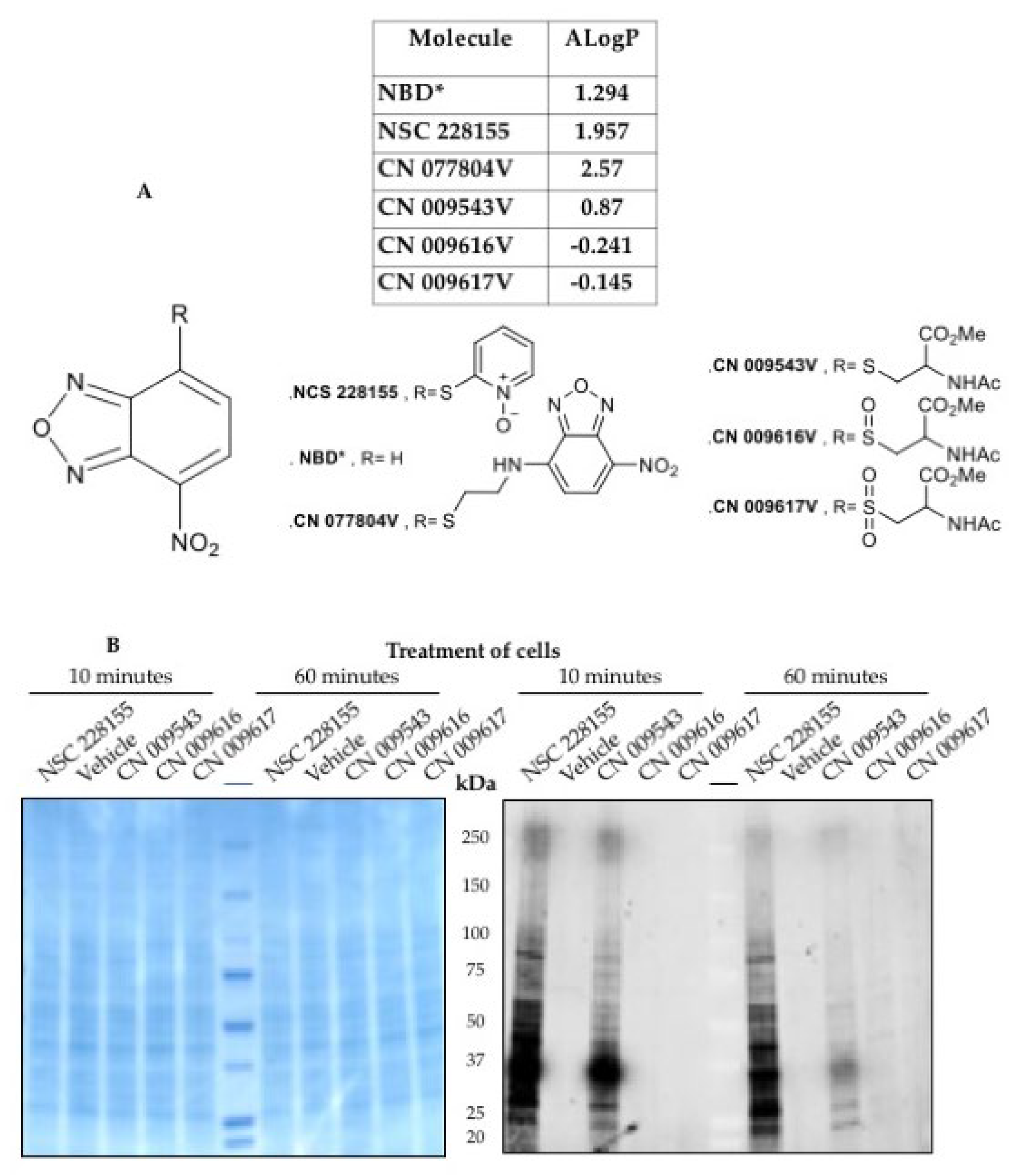
We took advantage of the fluorescence of the NBD structure to determine how its derivatives enter cells. Signals of different colors for the compound, plasma, and nucleus were detected using fluorescence microscopy and recorded as curves of different colors. NBD compounds have different abilities to penetrate cells [29] (Figure 3C). Compound NSC 228155 penetrates cell membranes and disperses in both the cytoplasmic and nuclear compartments, whereas compound CN 009616V cannot penetrate the lipid bilayer. A compound containing only the NBD scaffold severely damages the plasma membrane at the concentration used. This behavior is consistent with the lipophilicity of NBD compounds according to the calculated logP value (Figure 4A). The compound having a positive logP can enter cells and enhance tyrosine phosphorylation of EGFR in cells. In contrast, a compound with negative logP cannot cross the plasma membrane and therefore does not induce phosphorylation. The differential effects of NBD compounds on the cell can be important for subsequent understanding of the effect of hydrogen peroxide on the phosphorylation of many proteins.
A wide range of modern methods are used to detect ROS and their actions, including fluorescent [36] and chemiluminescent assays [37], chromatographic [38] and spectrophotometric assays [39,40], electrochemical biosensors [41] and electron paramagnetic resonance [42]. Small fluorescent molecules capable of generating H2O2 have proven important for the simultaneous detection of a large number of interacting partners in living cells.
We turned to the principle of immobilization of proteins on a nitrocellulose membrane with subsequent recognition of the reacting proteins by fluorescent antibodies [43,44]. However, in the method, known as “in-gel” (45), target proteins can be detected using fluorescent compounds that bind to proteins directly in the cell, without the need for a specific antibody binding step. The fluorescent signal emitted by bound proteins was recorded after 5-min and 60-min incubation of cells with 10 μM lipophilic NBD compounds (Figure 4B) [46]. It is pertinent to note that lipophilic compounds bind to many proteins having a molecular weight from 10 kDa to almost 300 kDa. The observed decrease in fluorescence intensity of bound proteins upon 60-min treatment may be due to degradation of the reacting proteins or dye inactivation.
Two lipophilic NBD compounds bind to many proteins with similar spectra, suggesting similar molecular weights of the bound proteins (Figure 4B). Interestingly, the lipophilic compounds NSC 228155 and CN 00943, which have different structures (Figure 4A and Figure 4B), exhibit the same interaction profile, probably with all proteins. This suggests that protein interaction occurs rather through the NBD core present in both compounds.
H2O2 interacts with various human cell proteins and regulates the expression of transcription factors through both activation and inactivation [47]. The described in-gel method appears attractive in future studies for the rapid and comparative assessment of many NBD-mediated interactions with proteins, such as members of protein tyrosine phosphatases, under different cell culture conditions.
2.3. Nrf2-ARE Redox Pathway
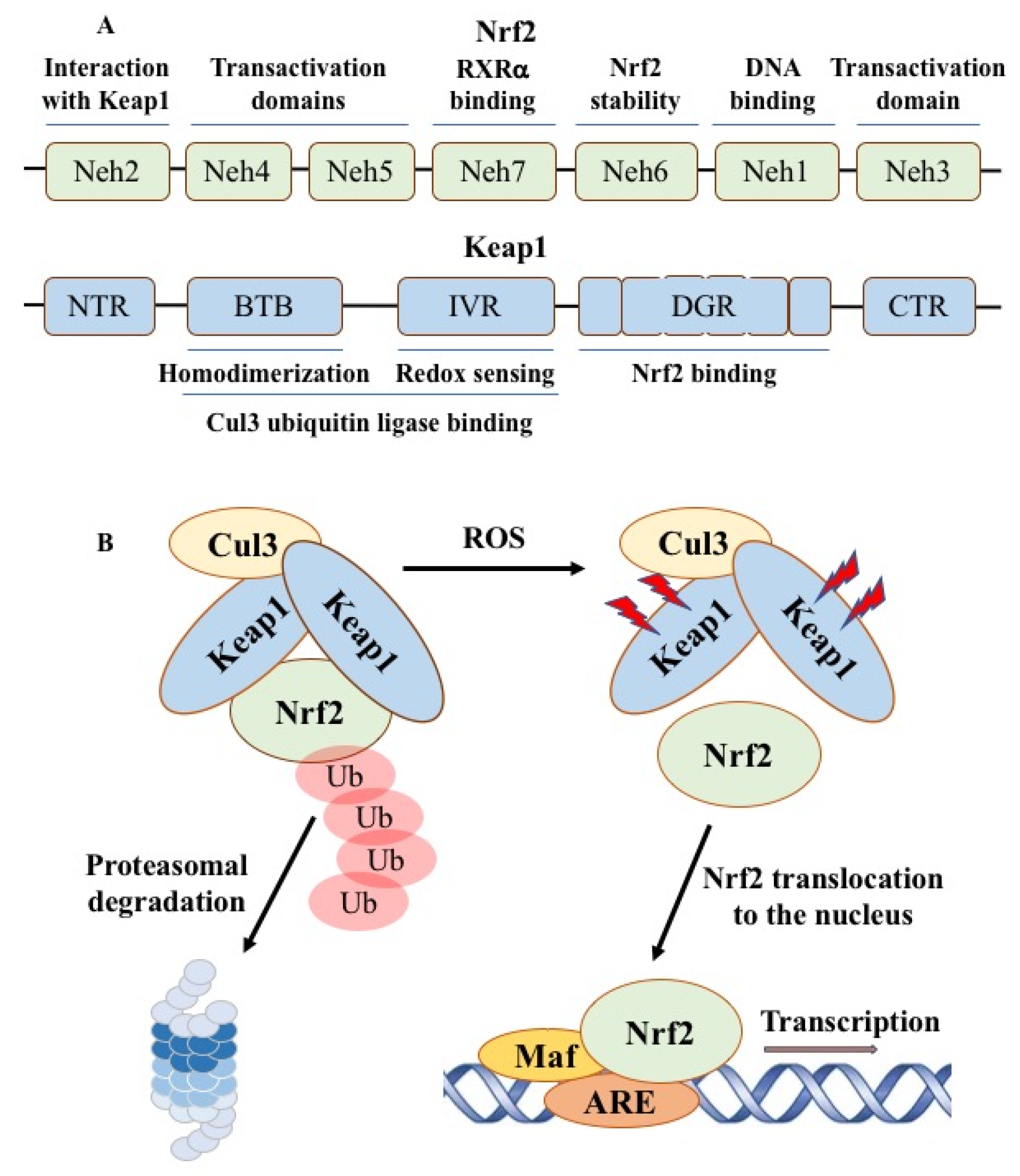
The cellular system for regulating ROS levels was discovered by isolating the transcription factor Nrf2 (nuclear factor E2-related factor 2), which allows the assessment of increased amounts of free radicals [48]. Constitutive activation of Nrf2 plays an important role in oxidative stress resistance, metabolic reprogramming, ferroptosis inhibition, and chemotherapy resistance of cancer cells, and is associated with poor prognosis and survival [49,50]. Nrf2 consists of seven Nrf2-ECH homology (Neh) cap “n” collar (CNC) domains (Figure 5). Recognition and binding of Nrf2 to sensitive genes is determined by the antioxidant response element EpRE/ARE, the activity of which is determined by the state of the redox-sensitive inhibitor Keap1 (Kelch-like ECH-associated protein 1), which acts as an adapter for the cullin E3 ubiquitin ligase [51]. With increasing oxidative stress, Keap1-reactive cysteine residues are oxidized, resulting in conformational changes that inhibit Nrf2 binding to Keap1 and allow the protein to escape proteolytic degradation.
Figure 4.
Structure and ALogP values of NBD compounds (A) and binding of NBD compounds to proteins in treated cell extracts detected by in-gel method (B). An image of proteins stained with Coomassie blue (left) and an image of many proteins detected on a nitrocellulose membrane in a fluorescence scan at Exλ 488 nm and Emλ 520 nm (right). The membrane was scanned using a Typhoon 9410 imaging system (Molecular Devices) equipped with filters of different wavelengths.
Figure 4.
Structure and ALogP values of NBD compounds (A) and binding of NBD compounds to proteins in treated cell extracts detected by in-gel method (B). An image of proteins stained with Coomassie blue (left) and an image of many proteins detected on a nitrocellulose membrane in a fluorescence scan at Exλ 488 nm and Emλ 520 nm (right). The membrane was scanned using a Typhoon 9410 imaging system (Molecular Devices) equipped with filters of different wavelengths.
With increasing oxidative stress, Keap1-reactive cysteine residues are oxidized, resulting in conformational changes that inhibit Nrf2 binding to KEAP1 and further movement of Nrf2 toward ubiquitination. Nrf2 determines the antioxidant response by binding to the ARE, a cis-acting regulatory element responsible for the expression of detoxification enzymes [52]. In the absence of oxidative stress, Nrf2 is ubiquitinated and degraded with a half-life of 30-60 min by the ubiquitin-proteasome system. Under conditions of oxidative stress caused by reactive electrophiles, toxins, or ARE inducers, the interaction between Nrf2 and Keap1 is disrupted and Nrf2 translocates into the nucleus, where it binds to small Maf proteins, increasing the rate of transcription of recognized genes. Thus, electrophilic compounds producing hydrogen permease covalently modify cysteine residues in the Keap1 protein and activate Nrf2, which triggers an antioxidant response and protects cells from oxidative stress.
A recent article describes the variety of beneficial effects of electrophilic compounds that interact with target proteins [53]. Nrf2 plays a critical role in maintaining cellular redox homeostasis and regulating cellular antioxidants such as glutathione and thioredoxin, as well as stimulating the expression of enzymes involved in NADPH and ROS regeneration, xenobiotic reduction and detoxification [54,55]. Nrf2 is the main regulator of cytoprotective processes, which can lead to the development and progression of human diseases, including cancer. Its expression closely correlates with drug resistance of tumour cells [56]. In colorectal cancer cells with Nrf2 knockdown, changes occurring in signalling pathways were associated with loss of ROS scavenging and detoxification potential [57]. In the absence of Nrf2, certain activated pathways such as MAPK, JNK and FOXO appear to reduce the deleterious effects of redox deficiency, and as knockdown cells become more sensitive to drugs, these pathways may be targeted to treat cancer. These observations do not exclude the possibility of activation of pathways other than Nrf2-ARE, which may contribute to a cytoprotective response against ROS and electrophiles [58].
Figure 5.
Domain structure of Nrf2 and Keap1 proteins and Nrf2 activation in response to ROS stress conditions. In the absence of stress, the Nrf2 protein complex is ubiquitinated by the Cul3 ubiquitin ligase and Nrf2 continuously degraded by the cellular proteasome. In response to ROS stress, Keap1 is inactivated, which leads to stabilization of Nrf2 and its passage into the nucleus and dimerization with small Maf proteins, followed by binding to the antioxidant response element ARE to activate transcription of various target proteins.
Figure 5.
Domain structure of Nrf2 and Keap1 proteins and Nrf2 activation in response to ROS stress conditions. In the absence of stress, the Nrf2 protein complex is ubiquitinated by the Cul3 ubiquitin ligase and Nrf2 continuously degraded by the cellular proteasome. In response to ROS stress, Keap1 is inactivated, which leads to stabilization of Nrf2 and its passage into the nucleus and dimerization with small Maf proteins, followed by binding to the antioxidant response element ARE to activate transcription of various target proteins.
Typically, the Nrf2 regulatory system takes about an hour to transcribe the Nrf2/Keap genes and translate the corresponding protein that recognizes the oxidant H2O2. The in-gel method quickly, in almost five minutes, detects binding to a large number of proteins that react differently to hydrogen peroxide. This means that the natural Nrf2 defence system, which takes longer to activate, appears to be unable to prevent H2O2 from binding to proteins and protect the human body from exposure to unwanted proteins.
Other stress-responsive transcription factors are also induced by the same types of reactive toxicants as Nrf2, indicating that the overall cellular response to oxidants also involves other pathways such as HSF1 and HIF1 [59]. Cellular stress perception and defence activity are regulated by these three oxidant-activated transcription factors HSF1, HIF1 and Nrf2, which control the heat shock response (unfolded proteins), hypoxia response and antioxidant response, respectively [48,60,61]. These three system responses create a multi-layered cellular defence consisting primarily of non-overlapping programs that mitigate the limitations of each response.
2.4. Therapeutic Opportunities by Targeting ROS-Producing Proteins
Human cellular responses to ROS are complex and dysregulation of ROS contributes to the development of cancer [62]. However, whether ROS can prevent or promote cancer development remains controversial.
The intracellular redox state is highly dynamic and tightly regulated by the coordinated actions of ROS generating and scavenging enzyme systems [63]. Human cells use a complex scavenging system in the cytoplasm, mitochondria and extracellular matrix, including superoxide dismutase (SOD), glutathione peroxidase, glutathione reductase, peroxiredoxin, thioredoxin and catalase, as well as other antioxidants that convert superoxide anions into water [64]. Endogenous ROS generated during inflammation can cause chemical damage to macromolecules, namely DNA, proteins and lipids. Damage to intracellular macromolecules leads to oxidative stress during cellular respiration or inflammation under aerobic conditions. In DNA, the guanine base is the most electron-rich and most prone to oxidation, which is converted into several oxidation products, the main one being 8-oxo-7,8-dihydroguanine. When DNA is damaged by free radicals, the amount of 8-oxo-7,8-dihydroguanine increases, which has made it possible to use this molecule as a biomarker of oxidative stress [65]. Oxidative damage to proteins affects their activity, leading to increased target proteolysis and dysfunction of signaling pathways [66]. As a result of lipid peroxidation by ROS, the resulting lipid radicals, alkanes and isopropanes can affect the course of the pathological process, especially in diabetes and neurodegenerative diseases [67]. In these studies, the addition of H2O2 to the in vitro culture medium is commonly used to induce cellular oxidative stress.
Biological thiols, such as glutathione and N-acetylcysteine, play an important role in maintaining redox homeostasis by acting as antioxidants and scavenging free radicals [68]. Glutathione is a tripeptide antioxidant composed of glutamic acid, cysteine and glycine and is part of the endogenous defense against ROS. N-acetylcysteine provides the cysteine structure of glutathione and improves the intracellular content of the -SH group. In healthy cells and tissues, more than 90% of total glutathione levels are in the reduced thiol form, with the remainder in the oxidized disulfide form. In the global redox homeostasis of the cell, the level of disulfide-glutathione is maintained at a low level, while the reduced thiol-glutathione remains high. Therefore, an increased glutamine disulfide/thiol ratio can be considered an indicator of oxidative stress [69,70].
An antioxidant is a relatively stable low-molecular compound that can donate an electron to a free radical and neutralize it, thereby reducing the potential of the radical to harm the human body. Antioxidants act through various mechanisms as radical scavengers, hydrogen donors, electron donors, peroxide decomposers, singlet oxygen quenchers, enzyme inhibitors, synergists, or metal chelators [67]. Activation of the human Nrf2 defense system can be provided by curcumin (from the spice turmeric), resveratrol (from grapes), sulforaphane (from broccoli), and antioxidants found in other plant foods [71,72]. In contrast, the defense system can be inhibited by other natural products such as apigenin, luteolin, chrysin, which have the potential to counteract oxidative stress. In addition to natural sources of antioxidants, it has been demonstrated that chemically synthesized molecules can also exhibit similar activity.
The inflammatory microenvironment in disease is characterized by elevated levels of ROS and reactive nitrogen species (RNS), leading to oxidative stress. Antioxidant enzymes function as peroxide sensors and are emerging as key regulators of the many pathways that respond to hydrogen peroxide and therefore slow the progression of cancer, neurodegenerative diseases and other pathologies [73]. Decreased redox buffering capacity, resulting in antioxidant-oxidative imbalance, leads to an inflammatory response in cells and the progression of cancer, diabetes, and neurodegenerative diseases, including major depressive disorders [74]. Natural foods are considered potential antioxidants if they counteract the effects of oxidative stress in the pathogenesis of various diseases and aging. For example, bee products such as honey, propolis and royal jelly act as radical scavengers and are considered natural antioxidants [75,76]. Therefore, evaluation of the antioxidant activity of products is necessary to select the best samples that meet the requirements of medical use.
Synthetic antioxidants are chemicals widely used as additives to plastics, rubbers, cables and other polymeric materials to prevent their oxidative degradation [77,78]. Various exogenous substances, including pollutants, food additives, medications, or lifestyle factors, trigger ROS production [79]. Smoking continues to be the leading risk factor for cancer worldwide, as tobacco smoke contains thousands of chemicals, including more than 70 carcinogens [80]. Smoking causes chronic and progressive inflammation, which is a key factor in the pathophysiological progression of cancer and other diseases [81]. The industrial production of synthetic antioxidants leads to large-scale emissions into the environment and climate change, which creates devastating consequences for the plant world [82].
Chemical compounds in pollutants, when the content of reactive molecules is hundreds of times higher than permissible doses, can quickly penetrate cells and have a detrimental effect on the human body. In this regard, multifaceted protein microarrays, developed primarily for drug discovery purposes, are promising tools for cost-effective assessment of the biological reactivity of vast numbers of synthetic molecules [83]. Volatile lipophilic or non-lipophilic electrophilic ROS in atmospheric emissions can trigger the destruction of the ozone layer with subsequent undesirable effects on the biological systems of our planet. Undoubtedly, the study of electrophilic ROS is an integral part of the global challenge in our efforts to prevent the progression of cancer and other diseases. Today humanity is faced with an urgent need to include the products of large-scale transformation in environmental standards for nature protection in contaminated areas.
Hydrogen peroxide in low concentrations is used as an antiseptic for minor cuts. The US Food and Drug Administration (FDA) classifies hydrogen peroxide as GRAS - generally recognized as safe - for humans at low doses. However, H2O2 causes irritation, burning and blistering of the skin. The FDA does not recognize it as a dietary supplement. In February 2022, Medical Faculty Associates reported that the claim that topical application of hydrogen peroxide to the skin can treat cancer is unfounded [https://gwdocs.com/news/fact–check–false–claim–rubbing–hydrogen–peroxide–skin–treats–cancer–usatodaycom].
The US federal government estimated that in 2023, nearly 2 million Americans would be diagnosed with cancer (other than nonmelanoma skin cancer) and more than 600,000 people would die from the disease [https://www.aacr.org/patients–caregivers/awareness–months/national–cancer–prevention]. More than 40% of these cases and almost half of the deaths can be attributed to preventable causes - smoking, excessive sun exposure, excess body weight and lack of physical activity. That is, the potential role of ROS, including hydrogen peroxide, may be significant in the actual cause of cancer. Suggested measures such as quitting smoking and protecting your skin from the sun may help reduce the risk of cancer caused by ROS.
The binding of hydrogen peroxide to a targeted protein potentially stimulates the chemical molecule to stop the disease process. A number of laboratories were forced to stop research on the creation of new antioxidant drugs against a number of diseases, including due to the lack of a positive effect on proteins of therapeutic importance, such as glutathione S-transferase, c-Myc oncogene and HIV intergrase [84,85,86,87].
The powerful antioxidant edaravone was originally developed as a neuroprotective agent for acute cerebral infarction and neurological pathologies [88]. But after trials, it became attractive only for the treatment of amyotrophic lateral sclerosis. Data from animal models indicate its possible safety in humans [89]. However, the lack of clinical trial data does not allow us to talk about the real therapeutic value of edaravone.
Reliable drugs that stop the progression of oncological, neurological, allergic and other disease models under the influence of non-reactive ROS-producing compounds have not been described. The failure to effectively modulate hydrogen permease activity with chemical compounds in laboratory and preclinical trials suggests that the problem is more complex than previously thought. It can be assumed that the stumbling block to successful therapy is not only the acquisition of resistant mutations in the gene encoding SOD, but also the ineffectiveness of the treatment strategy itself, which is observed in the fight against H2O2-associated cancer.
On what basis do we build this consideration? Considering the data presented in this review article, the following factors can be noted. First, hydrogen peroxide and the H2O2-producing NBD compounds affect a huge number of different proteins by directly binding to them. Secondly, among the target proteins there are proteins with opposite activities due to mutations or other intracellular reasons. Third, hydrogen peroxide plays a dual and opposite role in its action on proteins, activating some proteins and inactivating other proteins. Fourth, it cannot be excluded that the direction of influence on the target process changes over time when the activity of one of the two proteins modulates with changes in environmental conditions. Fifth, activation of the natural Nrf2 defense system in aerobic mammals requires more time to bind to proteins than the much shorter binding time of the non-reactive H2O2 molecule. Therefore, actual protection of diseased cells may not be possible due to the rapid binding of hydrogen peroxide to the target protein.
These observations lead to the important pharmacological and medical conclusion that superoxide dismutase and possibly other H2O2-producing proteins cannot be inhibited for therapeutic purposes. Unfortunately, the nature of these proteins does not provide a suitable basis for the development of chemical therapies for diseases caused or associated with ROS.
3. Conclusions
The evolution of aerobic organisms on Earth was associated with a constant increase in oxygen content in the atmosphere. Free radicals, such as reactive oxygen and reactive nitrogen species (RNS), readily react with other chemical and biological entities in living cells. Various defense mechanisms have evolved to promote the diversity of life in the oxygen world on this planet [90].
It is appropriate to emphasize that reactive oxygen species play an important role not only in biological processes, but also in the military field [www.youtube.com/watch?v=jnyHp1LJKQk]. Let’s not forget that two free radicals - the superoxide radical and the hydroxyl radical - have an extremely rapid ability to destroy biological systems, leading to human death.
In the absence of progress in the development of drugs that counteract the negative effects of ROS and RNS, the question arises about the legitimacy of considering chemical compounds as reliable therapeutic agents that affect superoxide dismutase and other proteins with similar functional activity. Reducing exposure can not only have a positive impact on efforts to reduce the burden of disease, but also synergistically improve human health.
Given the impossibility of developing drugs against superoxide dismutase and its related proteins, we changed our strategy to create chemical compounds that can inhibit cancer cells caused by EGFR activation [92,93]. Understanding the concomitant effects of a therapeutic agent on many different target proteins is an important strategy for preventing side effects.
Author Contributions
V.S. - preparation of the initial draft, final manuscript and figures, supervision; R.A.d.S. - data analysis, preparation of figures. The authors have agreed to the published version of the manuscript.
Funding
This review received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request to the authors.
Acknowledgments
The authors express their gratitude to the colleagues who took part in the experiments and provided their professional knowledge.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Auten, R.; Davis, J. Oxygen toxicity and reactive oxygen species: The devil is in the details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Miller, A.F. Superoxide dismutases: ancient enzymes and new insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Fridovich, I. Superoxide anion radical (O2-. ), superoxide dismutases, and related matters. J. Biol. Chem. 1997, 272, 18515–18517. [Google Scholar] [PubMed]
- Casas, A.I.; Nogales, C.; Mucke, H.A.M.; Petraina, A.; Cuadrado, A.; Rojo, A.I.; Ghezzi, P.; Jaquet, V.; Augsburger, F.; Dufrasne, F.; Soubhye, J.; Deshwal, S.; Di Sante, M.; Kaludercic, N.; Di Lisa, F.; Schmidt, H.H.H.W. On the clinical pharmacology of reactive oxygen species. Pharmacol. Rev. 2020, 72(4), 801–828. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The role of hydrogen peroxide in redox-dependent signaling: homeostatic and pathological responses in mammalian cells. Cells 2018, 7(10), 156. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, D.; Abbruzzese, J.L.; Lu, W. Measurement of Reactive Oxygen Species by Fluorescent Probes in Pancreatic Cancer Cells. In: Su, G. (eds) Pancreatic Cancer. Methods in Molecular Biology. (2019), 1882. Humana Press, New York, NY.
- Loschen, G.; Azzi, A.; Richter, C.; Flohé, L. Superoxide radicals as precursors of mitochondrial hydrogen peroxide. FEBS Lett. 1974, 42(1), 68–72. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, P. , Jaquet V., Marcucci F., Schmidt H.H.H.W. The oxidative stress theory of disease: levels of evidence and epistemological aspects. Br. J. Pharmacol. 2017, 174(12), 1784-1796. [CrossRef]
- Lemmon M., A.; Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef]
- Paul, M. D.; Grubb, H.N.; Hristova, K. Quantifying the strength of heterointeractions among receptor tyrosine kinases from different subfamilies: Implications for cell signaling. J. Biol. Chem. 2020, 295, 9917–9933. [Google Scholar] [CrossRef]
- Jorissen, R.N.; Walker, F.; Pouliot, N.; Garrett, T.P.J.; Ward, C.W.; Burgess, A.W. Epidermal Growth Factor Receptor: Mechanisms of Activation and Signalling. Exp. Cell Res. 2003, 284, 31–53. [Google Scholar] [CrossRef]
- Cohen, S. Origins of growth factors: NGF and EGF. J. Biol. Chem. 2008, 283(49), 33793–33797. [Google Scholar] [CrossRef] [PubMed]
- Jura, N.; Zhang, X.; Endres, N.F.; Seeliger, M.A.; Schindler, T.; Kuriyan, J. Catalytic control in the EGF receptor and its connection to general kinase regulatory mechanisms. Mol. Cell. 2011, 42, 9–22. [Google Scholar] [CrossRef]
- Gamou, S.; Shimizu, N. Hydrogen peroxide preferentially enhances the tyrosine phosphorylation of epidermal growth factor receptor. FEBS Lett. 1995, 357, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S. , Kang, S. W.; Seo, M.S.; Baines, I.C.; Tekle, E.; Chock, P.B.; Rhee, S.G. Epidermal growth factor (EGF)-induced generation of hydrogen peroxide. Role in EGF receptor-mediated tyrosine phosphorylation. J. Biol. Chem. 1997, 272, 217–221. [Google Scholar]
- Zhang, J.; Yang, P.L.; Gray, N.S. Targeting cancer with small molecule kinase inhibitors. Nat. Rev. Cancer 2009, 9(1), 28–39. [Google Scholar] [CrossRef]
- Paulsen, C.E.; Truong, T.H.; Garcia, F.J.; Homann, A.; Gupta, V.; Leonard, S.E.; Carroll, K.S. Peroxide-dependent sulfenylation of the EGFR catalytic site enhances kinase activity. Nat. Chem. Biol. 2011, 8, 57–64. [Google Scholar] [CrossRef]
- Truong, T.H.; Carroll, K.S. Redox regulation of epidermal growth factor receptor signaling through cysteine oxidation. Biochemistry 2012, 51, 9954–9965. [Google Scholar] [CrossRef]
- Ferrari, E.; Tinti, M.; Costa, S.; Corallino, S.; Nardozza, A.P.; Chatraryamontri, A.; Ceol, A.; Cesareni, G.; Castagnoli, L. Identification of new substrates of the protein-tyrosine phosphatase PTP1B by Bayesian integration of proteome evidence. J. Biol. Chem. 2011, 286(6), 4173–4185. [Google Scholar] [CrossRef]
- Lee, S.R. , Kwon, K.S., Kim, S.R., Rhee, S.G. Reversible inactivation of protein-tyrosine phosphatase 1B in A431 cells stimulated with epidermal growth factor. J. Biol. Chem. 1998, 273(25), 15366-15372. [CrossRef]
- Salmeen, A.; Andersen, J.N. , Myers, M.P., Meng, T.C.; Hinks, J.A.; Tonks, N.K.; Barford, D. Redox regulation of protein tyrosine phosphatase 1B involves a sulphenyl-amide intermediate. Nature 2003, 423(6941):769-773. [CrossRef]
- van Montfort, R.L.; Congreve, M.; Tisi, D.; Carr, R. ; Jhoti, H. Oxidation state of the active-site cysteine in protein tyrosine phosphatase 1B. Nature 2003, 423(6941), 773-777. [CrossRef]
- Schwartz, P.A.; Kuzmic, P., Solowiej, J.; Bergqvist, S.; Bolanos, B.; Almaden, C.; Nagata, A.; Ryan, K.; Feng, J.; Dalvie, D.; Kath, J.C.; Xu, M.; Wani, R.; Murray, B.W. Covalent EGFR inhibitor analysis reveals importance of reversible interactions to potency and mechanisms of drug resistance. Proc. Natl. Acad. Sci. U S A 2014, 111(1), 173–178. [CrossRef]
- Sakanyan, V.; Angelini, M.; Le Béchec, M.; Lecocq, M.F.; Benaiteau, F.; Rousseau, B.; Gyulkhandanyan, A.; Gyulkhandanyan, L.; Logé, C.; Reiter, E.; Roussakis, C.; Fleury, F. Screening and discovery of nitro-benzoxadiazole compounds activating epidermal growth factor receptor (EGFR) in cancer cells. Sci. Rep. 2014, 4, 3977. [Google Scholar] [CrossRef]
- Toyo’oka, T. Development of Benzofurazan−bearing Fluorescence Labeling Reagents for separation and detection in high−performance liquid chromatography. Chromatography, 2012, 33, 1–17. [Google Scholar] [CrossRef]
- Blair, J. A.; Rauh, D.; Kung, C.; Yun, C.H.; Fan, Q.W.; Rode, H.; Zhang, C.; Eck, M.J.; Weiss, W.A.; Shokat, K. M. Nat. Chem. Biol. 2007, 3, 229–238. [Google Scholar]
- Heyne, B. ; Beddie, C; Scaiano, J. C. Synthesis and characterization of a new fluorescent probe for reactive oxygen species. Org. Biomol. Chem. 2007, 5, 1454–1458. [Google Scholar]
- Sakanyan, V.; Hulin, P.; Alves de Sousa, R.; Silva, V.; Hambardzumyan, A.; Nedellec, S.; Tomasoni, C.; Logé, C.; Pineau, C.; Roussakis, C.; Fleury, F.; Artaud, I. Activation of EGFR by small compounds through coupling the generation of hydrogen peroxide to stable dimerization of Cu/Zn SOD1. Sci. Rep. 2016, 6, 21088. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.A.O.; Lafont, F.; Benhelli-Mokrani, H.; LeBreton, M.; Hulin, P.; Chabot, T.; Paris, F.; Sakanyan, V.; Fleury, F. Rapid diminution in the level and activity of DNA-dependent protein kinase in cancer cells by a reactive nitro-benzoxadiazole compound. Int. J. Mol. Sci. 2016, 17(5), 703. [Google Scholar] [CrossRef] [PubMed]
- Burma, S.; Chen, B.P.; Chen, D.J. Role of non-homologous end joining (NHEJ) in maintaining genomic integrity. DNA Repair 2006, 5, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Groeger, A.L.; Freeman, B.A. Signaling actions of electrophiles: anti-inflammatory therapeutic candidates. Mol. Interv. 2010, 10(1), 39–50. [Google Scholar] [CrossRef] [PubMed]
- Schülke, K.H.; Ospina, F.; Hörnschemeyer, K.; Gergel, S.; Hammer, S.C. Substrate profiling of anion methyltransferases for promiscuous synthesis of S-Adenosylmethionine analogs from haloalkanes. Chembiochem. 2022, 23(4), e202100632. [Google Scholar] [CrossRef]
- Edwards, D.R.; Lohman, D.C.; Wolfenden, R. Catalytic proficiency: the extreme case of S-O cleaving sulfatases. J. Am. Chem. Soc. 2012, 134(1), 525–531. [Google Scholar] [CrossRef]
- Kenakin, T.; Miller, L.J. Seven transmembrane receptors as shapeshifting proteins: the impact of allosteric modulation and functional selectivity on new drug discovery. Pharmacol. Rev. 2010, 62, 265–304. [Google Scholar] [CrossRef]
- Zhao, H.; Joseph, J.; Fales, H. M.; Sokoloski, E. A.; Levine, R. L.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc. Natl. Acad. Sci. U S A 2005, 102, 5727–5732. [Google Scholar] [CrossRef] [PubMed]
- Kojima, R; Takakura, H. Kamiya, M.; Kobayashi, E.; Komatsu, T.; Ueno, T.; Terai, T.: Hanaoka, K.; Nagano, T.; Urano, Y. Development of a sensitive bioluminogenic probe for imaging highly reactive oxygen species in living rats. Angew. Chem. Int. Ed. Engl. 2015, 54, 14768–14771.
- Tsamesidis, I.; Egwu, C.O.; Pério, P.; Augereau, J.M.; Benoit-Vical, F.; Reybier, K. An LC-MS Assay to measure superoxide radicals and hydrogen peroxide in the blood system. Metabolites 2020, 10(5), 175. [Google Scholar] [CrossRef] [PubMed]
- Melov, S.; Coskun, P.; Patel, M.; Tuinstra, R.; Cottrell, B.; Jun, A.S.; Zastawny, T.H.; Dizdaroglu, M.; Goodman, S.I.; Huang, T.T.; Miziorko, H.; Epstein, C.J.; Wallace, D.C. Mitochondrial disease in superoxide dismutase 2 mutant mice. Proc. Natl. Acad. Sci. U S A 1999, 96, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ge, W.; Martínez-Jarquín, S.; He, Y.; Wu, R.; Stoffel, M.; Zenobi, R. Mass spectrometry reveals high levels of hydrogen peroxide in pancreatic cancer cells. Angew. Chem. Int. Ed. Engl. 2023, 62(19), e202213703. [Google Scholar] [CrossRef] [PubMed]
- Beissenhirtz, M.K.; Scheller, F.W.; Lisdat, F. A superoxide sensor based on a multilayer cytochrome c electrode. Anal. Chem. 2004, 76, 4665–71. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Qiu, T.; Yang-Ping, L.; Yang, L. Superoxide anion radical generation in the NaOH/H(2)O(2)/Fe(III) system: a spin trapping ESR study. Magn. Reson. Chem. 2006, 44(1), 38–44. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. U S A 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Burnette, W.N. "Western blotting": electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Johnson, I.; Spence, M. The Molecular probes handbook: A guide to fluorescent probes and labeling technologies. Life Technologies Corporation. 2010.
- Sakanyan, V.; Benaiteau, F.; Alves de Sousa, R.; Pineau, C.; Artaud, I. Straightforward detection of reactive compound binding to multiple proteins in cancer cells: Towards a better understanding of electrophilic stress. Ann. Clin. Exp. Metabol. 2016, 1, 1006. [Google Scholar]
- Marinho, H.S.; Real, C.; Zirn, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. U S A 1994, 91(21), 9926–9930. [Google Scholar] [CrossRef]
- Ohta, T.; Iijima, K.; Miyamoto, M.; Nakahara, I.; Tanaka, H.; Ohtsuji, M.; Suzuki, T.; Kobayashi, A.; Yokota, J.; Sakiyama, T.; Shibata, T.; Yamamoto, M.; Hirohashi, S. Loss of Keap1 function activates Nrf2 and provides advantages for lung cancer cell growth. Cancer Res. 2008, 68, 1303−1309. [Google Scholar] [CrossRef]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D. D. NRF2 and the hallmarks of cancer. Cancer Cell 2018, 34, 21−43. [Google Scholar] [CrossRef]
- Joshi, G.; Johnson, J.A. The Nrf2-ARE pathway: a valuable therapeutic target for the treatment of neurodegenerative diseases. Recent Pat. CNS Drug Discov. 2012, 7(3), 218–229. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by Nrf2. Antioxid. Redox Signal. 2018, 29(17), 1727–1745. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Bustamante Munguira, E.; Juan, C.A.; Plou, F.J.; Pérez, L.E. Electrophilic compounds in the human diet and their role in the induction of the transcription factor NRF2. Int. J. Mol. Sci. 2024, 25, 3521. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. U S A 2018, 115(23), 5839–5848. [Google Scholar] [CrossRef]
- Unoki, T.; Akiyama, M.; Kumagai, Y. Nrf2 activation and its coordination with the protective defense systems in response to electrophilic stress. Int. J. Mol. Sci. 2020, 21, 545. [Google Scholar] [CrossRef]
- Dempke, W.C.M.; Reck, M. KEAP1/NRF2 (NFE2L2) mutations in NSCLC - Fuel for a superresistant phenotype? Lung Cancer 2021, 159, 10−17. [Google Scholar] [CrossRef]
- Cheraghi, O.; Dabirmanesh, B.; Ghazi, F.; Amanlou, M.; Atabakhshi-Kashi, M.; Fathollahi, Y.; Khajeh, K. The effect of Nrf2 deletion on the proteomic signature in a human colorectal cancer cell line. BMC Cancer 2022, 22(1), 979. [Google Scholar] [CrossRef]
- Bono, S.; Feligioni, M.; Corbo, M. Impaired antioxidant KEAP1-NRF2 system in amyotrophic lateral sclerosis: NRF2 activation as a potential therapeutic strategy. Mol. Neurodegener. 2021, 16(1), 71. [Google Scholar] [CrossRef]
- Cyran, A.M.; Zhitkovich, A. HIF1, HSF1, and NRF2: Oxidant-responsive trio raising cellular defenses and engaging immune system. Chem. Res. Toxicol. 2022, 35(10), 1690–1700. [Google Scholar] [CrossRef]
- Ahn, S.G; Thiele, D.J. Redox regulation of mammalian heat shock factor 1 is essential for Hsp gene activation and protection from stress. Genes Dev. 2003, 17(4), 516–528. [Google Scholar] [CrossRef]
- Huang, L.E.; Arany, Z.; Livingston, D.M.; Bunn, H.F. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its alpha subunit. J Biol Chem. 1996, 271(50), 32253–32259. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell 2020, 38(2), 167–197. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2(5), e1600200. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: the bright side of the moon. Exp. Mol. Med. 2020, 52(2), 192–203. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nishigori, C.; Tanaka, T.; Uchida, K.; Nikaido, O.; Osawa, T.; Hiai, H.; Imamura, S.; Toyokuni, S. 8-hydroxy-2'-deoxyguanosine is increased in epidermal cells of hairless mice after chronic ultraviolet B exposure. J. Invest. Dermatol. 1996, 107(5), 733–737. [Google Scholar] [CrossRef] [PubMed]
- Lonkar, P.; Dedon, P.C. Reactive species and DNA damage in chronic inflammation: reconciling chemical mechanisms and biological fates. Int. J. Cancer 2011, 128, 1999−2009. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4(8), 118–126. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48 (2), 158−167. [CrossRef]
- Niki, E.; Tsuchiya, J.; Tanimura, R.; Kamiya, Y. Regeneration of vitamin E from a chromanoxyl radical by glutathione and vitamin C. Chem. Lett. Jpn. 1982, 11 (6), 789−792. [CrossRef]
- Gropper, S.S.; Smith, J.L.; Carr, T.P. Advanced nutrition and human metabolism, 7th Ed, Cengage Learning Inc., Wadsworth, 2017.
- Ashrafizadeh, M. ; Ahmadi, Z;; Mohammadinejad, R.; Farkhondeh, T.; Samarghandian, S. Curcumin activates the Nrf2 pathway and induces cellular protection against oxidative injury. Curr. Mol. Med. 2020, 20(2), 116-133. [CrossRef]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane and other nutrigenomic Nrf2 activators: Can the clinician's expectation be matched by the reality? Oxid. Med. Cell. Longev. 2016, 2016, 7857186. [Google Scholar] [CrossRef] [PubMed]
- Veal, E.A; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell. 2007, 26(1), 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Iordache, F.; Stanca, L.; Predoi, G.; Serban, A.I. Oxidative stress mitigation by antioxidants - an overview on their chemistry and influences on health status. Eur. J. Med. Chem. 2021, 209, 112891. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Yang, L.; Huang, Y.; Tao, L.; Chen, D. ′′Novel′′ synthetic antioxidants in house dust from multiple locations in the Asia-Pacific region and the United States. Environ. Sci. Technol. 2021, 55 (13), 8675−8682.
- Gong, X.; Zhang, W.; Zhang, S.; Wang, Y.; Zhang, X.; Lu, Y.; Sun, H.; Wang, L. Organophosphite antioxidants in mulch films are important sources of organophosphate pollutants in farmlands. Environ. Sci. Technol., 2021, 55(11), 7398-7406. [CrossRef]
- Liu, R.; Mabury, S.A. Synthetic phenolic antioxidants: A review of environmental occurrence, fate, human exposure, and toxicity. Environ. Sci. Technol. 2020, 54 (19), 11706−11719. [CrossRef]
- Liang, B.; Li, J.; Du, B.; Pan, Z.; Liu, L.-Y.; Zeng, L. E-Waste recycling emits large quantities of emerging aromatic amines and organophosphites: A poorly recognized source for another two classes of synthetic antioxidants. Environ. Sci. Technol. Lett. 2022, 9, 625−631. [Google Scholar] [CrossRef]
- Tian, Z.; Zhao, H.; Peter, K.T.; Gonzalez, M.; Wetzel, J.; Wu, C.; Hu, X.; Prat, J.; Mudrock, E.; Hettinger, R.; Cortina, A.E.; Biswas, R.G.; Kock, F.V.C.; Soong, R.; Jenne, A.; Du, B.; Hou, F.; He, H.; Lundeen, R.; Gilbreath, A.; Sutton, R.; Scholz, N.L.; Davis, J.W.; Dodd, M.C.; Simpson, A.; McIntyre, J.K.; Kolodziej, E.P. A ubiquitous tire rubber-derived chemical induces acute mortality in coho salmon. Science 2021, 371 (6525), 185−189. [CrossRef]
- Caliri, A.W.; Tommasi, S.; Besaratinia, A. Relationships among smoking, oxidative stress, inflammation, macromolecular damage, and cancer. Mutat. Res. Rev. Mutat. Res. 2021, 787, 108365. [Google Scholar] [CrossRef] [PubMed]
- Madani, A.; Alack, K.; Richter, M.J.; Kruger, K. Immune-regulating effects of exercise on cigarette smoke-induced inflammation. J. Inflamm. Res. 2018, 11, 155−167. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: generation, signaling, and defense mechanisms. Antioxidants (Basel) 2021, 10(2), 277. [Google Scholar] [CrossRef]
- Yeretssian, G.; Lecocq, M.; Lebon, G.; Hurst, H.C.; Sakanyan, V. Competition on nitrocellulose-immobilized antibody arrays: from bacterial protein binding assay to protein profiling in breast cancer cells. Mol. Cell. Proteomics 2005, 4(5), 605–617. [Google Scholar] [CrossRef]
- Sau, A.; Pellizzari Tregno, F.; Valentino, F.; Federici, G.; Caccuri, A.M. Glutathione transferases and development of new principles to overcome drug resistance. Arch. Biochem. Biophys. 2010, 500, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S; Prochownik, E. V. Small-molecule inhibitors of the Myc oncoprotein. Biochim. Biophys. Acta 2014, 1849, 525–543.
- Korolev, S.P.; Kondrashina, O.V.; Druzhilovsky, D.S.; Starosotnikov, A.M.; Dutov, M.D.; Bastrakov, M.A.; Dalinger, I.L.; Filimonov, D.A.; Shevelev, S.A.; Poroikov, V.V.; Agapkina, Y.Y.; Gottikh, M.B. Structural-functional analysis of 2,1,3-benzoxadiazoles and their N-oxides as HIV-1 integrase inhibitors. Acta Naturae. 2013, 5(1), 63–72. [Google Scholar] [CrossRef] [PubMed]
- Sakanyan, V. Reactive chemicals and electrophilic stress in cancer: A minireview. High Throughput. 2018, 7(2), 12. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Aoki, M.; Tsuji, S.; Itoyama, Y.; Sobue, G.; Togo, M.; Hamada, C.; Tanaka, M.; Akimoto, M., Nakamura, K.; et al.; Writing Group; Edaravone. (MCI-186) ALS 19 Study Group. Safety and efficacy of edaravone in well-defined patients with amyotrophic lateral sclerosis: a randomized, double-blind, placebo-controlled trial. Lancet Neurol. 2017, 16(7), 505–512. [CrossRef]
- Yamashita, T.; Abe, K. Update on antioxidant therapy with edaravone: expanding applications in neurodegenerative diseases. Int. J. Mol. Sci. 2024, 25(5), 2945. [Google Scholar] [CrossRef]
- Weinberg, F.; Ramnath, N.; Nagrath, D. Reactive oxygen species in the tumor microenvironment: An overview. Cancers 2019, 11(8), 1191. [CrossRef]
- Brauer, G. Handbook of preparative inorganic chemistry. Vol. 1. Translation editing by Reed F. (2nd ed.). New York: Academic Press. p. 140., 1963, ISBN 978-0-12-126601-1.
- Iradyan, M. , Iradyan, N., Hulin, P.; Hambardzumyan, A.; Gyulkhandanyan, A., Alves de Sousa, R., Hessani, A., Roussakis, C., Bollot, G., Bauvais, C.; Sakanyan, V. Targeting degradation of EGFR through the allosteric site leads to cancer cell detachment-promoted death. Cancers (Basel) 2019, 11(8), 1094. [CrossRef]
- Sakanyan, V.; Iradyan, N.; Alves de Sousa, R. Targeted strategies for degradation of key transmembrane proteins in cancer. BioTech (Basel) 2023, 12(3), 57. [Google Scholar] [CrossRef]
Figure 1.
Intracellular conversion between different kinds of ROS and action of antioxidant agents. SOD - superoxide dismutase, MPO - myeloperoxidase, GPx - glutathione peroxidase, Fe2+ is ferrous iron and participates in the Fenton reaction. ROS radicals can generate a new, highly reactive and unstable radical on a cellular partner. The antioxidant stabilizes this radical by donating an electron to make it less reactive.
Figure 1.
Intracellular conversion between different kinds of ROS and action of antioxidant agents. SOD - superoxide dismutase, MPO - myeloperoxidase, GPx - glutathione peroxidase, Fe2+ is ferrous iron and participates in the Fenton reaction. ROS radicals can generate a new, highly reactive and unstable radical on a cellular partner. The antioxidant stabilizes this radical by donating an electron to make it less reactive.
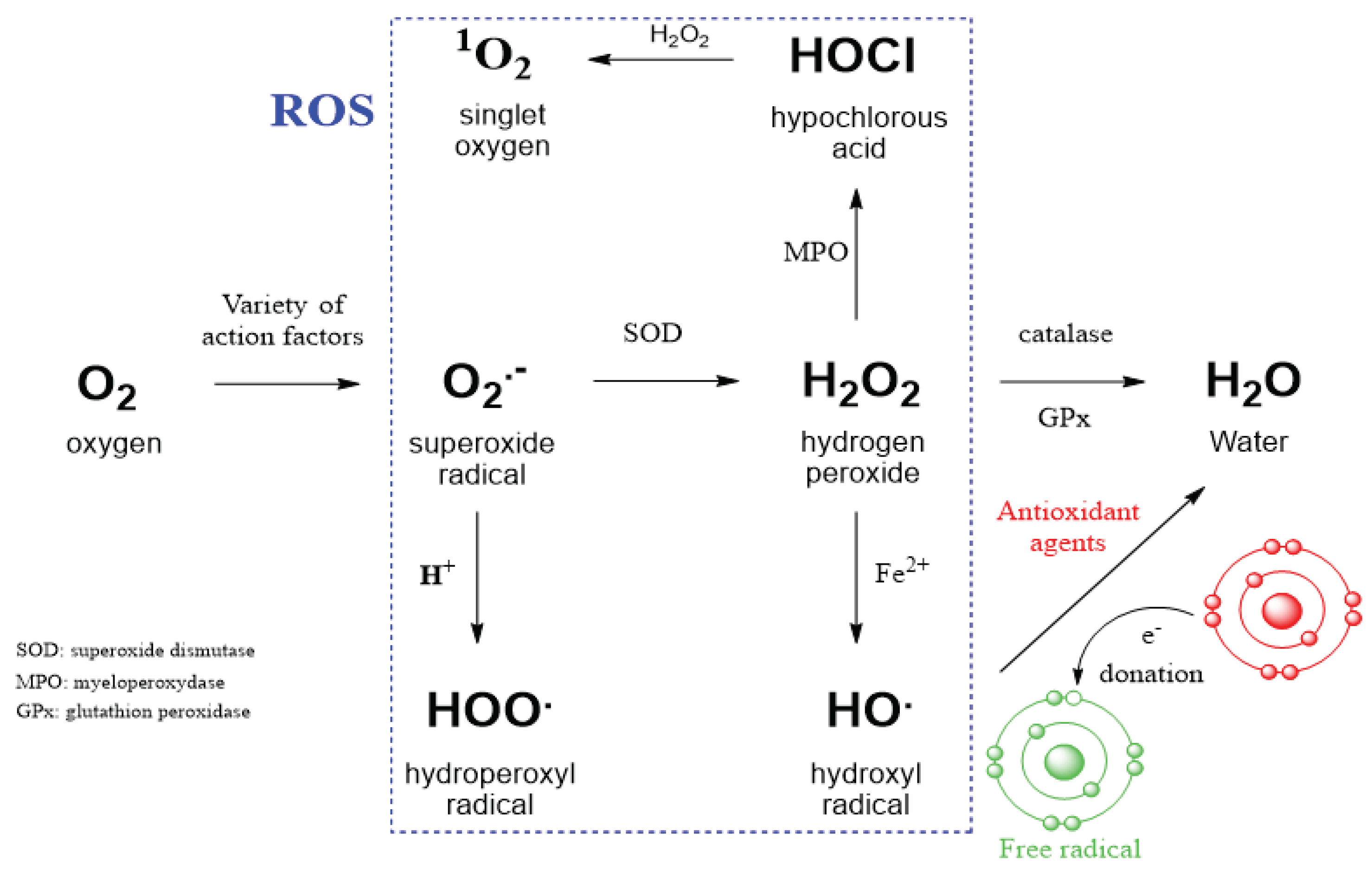
Figure 2.
Control of signaling pathways by EGFR. Extracellular binding of the cognate ligand EGF to EGFR results in dimerization of the receptor, followed by activation of the ATP binding site, phosphorylation of receptor tyrosine residues, and initiation of signaling pathways (three major pathways shown). Non-reactive H2O2 dimerizes EGFR in the absence of EGF in the growth medium and activates downstream signaling pathways.
Figure 2.
Control of signaling pathways by EGFR. Extracellular binding of the cognate ligand EGF to EGFR results in dimerization of the receptor, followed by activation of the ATP binding site, phosphorylation of receptor tyrosine residues, and initiation of signaling pathways (three major pathways shown). Non-reactive H2O2 dimerizes EGFR in the absence of EGF in the growth medium and activates downstream signaling pathways.
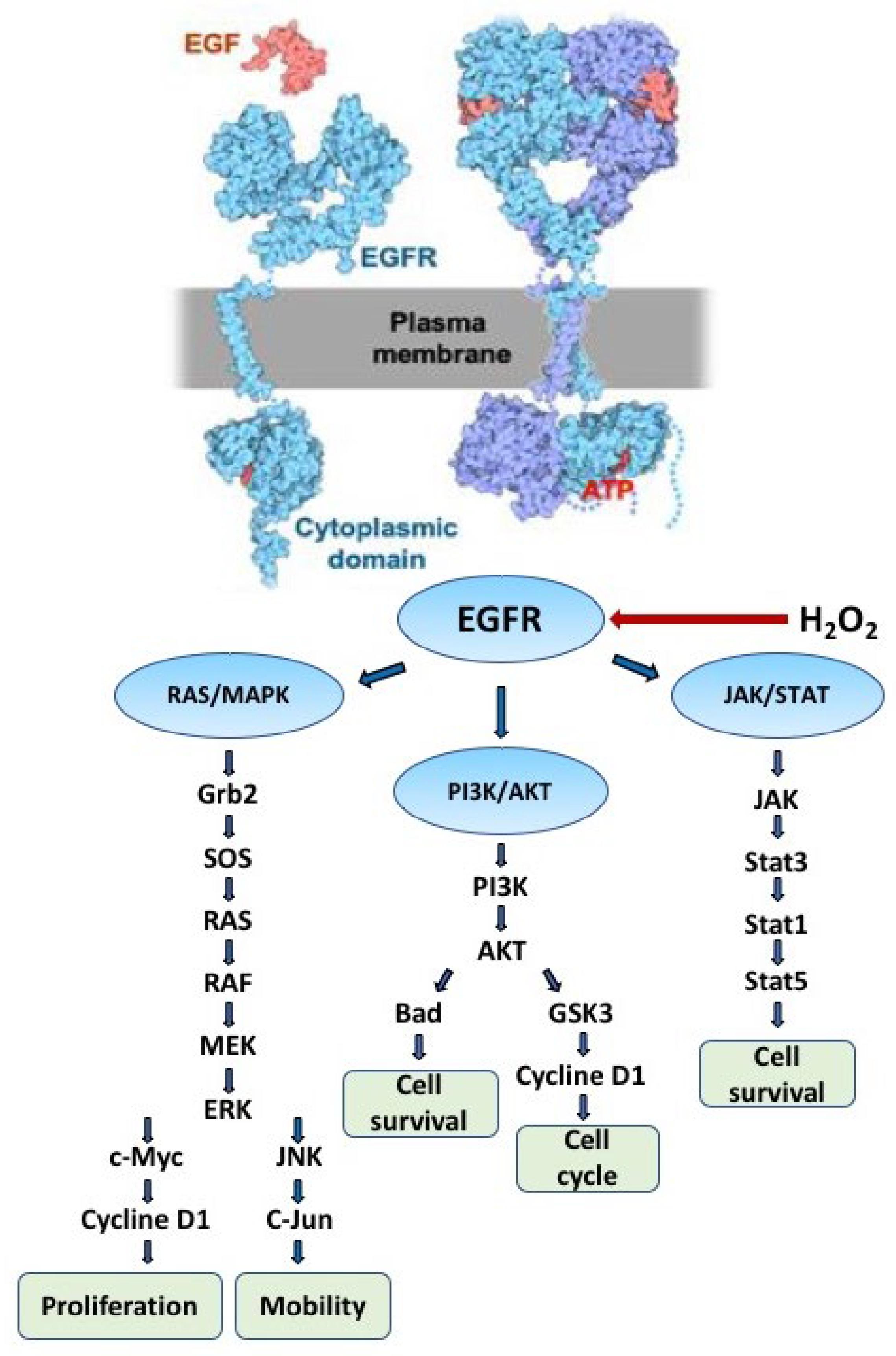
Figure 3.
Time-dependent activation of RTKs in breast cancer cells exposed to compound NSC 228155 (A) and compared with EGF (B), and distribution of NBD compounds detected by fluorescence microscopy (C). Cell images (left) and graphical representation of fluorescence (right) shown in representative cells exposed to NBD compounds. The wavy line shows the area being scanned; bar - 10 . Fluorescence of NBD compounds (emission 520 nm) is shown in green, plasma membrane (650 nm) in red, nucleus (420 nm) in blue.
Figure 3.
Time-dependent activation of RTKs in breast cancer cells exposed to compound NSC 228155 (A) and compared with EGF (B), and distribution of NBD compounds detected by fluorescence microscopy (C). Cell images (left) and graphical representation of fluorescence (right) shown in representative cells exposed to NBD compounds. The wavy line shows the area being scanned; bar - 10 . Fluorescence of NBD compounds (emission 520 nm) is shown in green, plasma membrane (650 nm) in red, nucleus (420 nm) in blue.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.


Dual Aquaporin and Sigma1 Receptor (DAS) Modulators: New Tools for Counteracting Oxidative Stress
Giorgia Pellavio
et al.
,
2020
A Tryptophan-based Assay Method to Search Regulatory Compounds for TCTP
Seri Jo
et al.
,
2020



MDPI Initiatives
Important Links
© 2024 MDPI (Basel, Switzerland) unless otherwise stated