
Preprint
Article
Preventive-Glycine Betaine Treatment Mitigate Spring and Summer Chilling Damage on Young Chestnut Trees under Climate Variability in Southern Chile
Altmetrics
Downloads
87
Views
43
Comments
0




Submitted:
07 May 2024
Posted:
08 May 2024
You are already at the latest version
Alerts
Abstract
Chestnut is an interesting productive alternative for farmers in Southern Chile. However, cli-matic variability with the occurrence of late spring and early summer frost have cause damages on young trees. Glycine‒betaine (GB) spraying on shoots of sensitive chestnut induce an increase in tolerance during subsequent chilling stress. Experiment was performed on three years-old trees (cultivar Cittá di Castello) conducted in a leader training system and sprayed with com-mercial GB (120 g hL‒1). In summer trees treated with GB showed a better recovery after the frost (‒2.34°C) occurrence for 3.5 h compared to the control. Whereas trees obtained a 28% greater growth of central axis than untreated control (January). The trunk diameter measured in January showed the highest value under GB (27.2mm) compared with the control (25.7 mm). The GB ap-plication increased by 50 and 47% highest fresh and dry weight respectively in leaves. Under GB trees reached a 28% higher net photosynthesis (AN) compared to the AN in control (11.93 µmol CO2 m‒2 s‒1). Moreover, achieved higher stomatal conductance and transpiration rate was 44 and 31% with GB compared to control trees respectively. Antioxidant performance was differentially affected by GB activating non-enzymatic mechanisms and caused a reduced lipid peroxidation. Glycine‒Betaine has become as an interesting agronomic tool that aimed to increase tolerance to environmental stresses such as low temperatures that cause damage to herbaceous shoots and leaves in young chestnut trees.
Keywords:
Subject: Biology and Life Sciences - Agricultural Science and Agronomy
1. Introduction
European chestnut (Castanea sativa L.) is an interesting productive alternative for farmers in Southern Chile. Currently, there are around 2,100 hectares (ha) covering this area with Italian and French cultivars (marroni) characterized by their high quality for both fresh consumption and industrialization. Most of the orchards are planted in foothills areas dominated by acid soils (pH<5.5) derived from volcanic ashes, that are characterized by good drainage and high organic matter contents. However, in recent years, climate variability with the occurrence of late spring and early summer frost causing severe damage on new shoots and leaves of young trees, triggering a series of morphological, physiological and biochemical changes particularly in the pre-Andean areas with a higher probability of frost occurrence (Figure 1A). As stressor factor, frost affects the adaptation of young chestnut orchards and consequently their projected productivity, which demands greater monetary costs and agronomical efforts.
There is evidence that it is possible to mitigate plant damage caused by low temperatures, during vegetative development, particularly on new tissues with osmoprotectant compounds, such as salicylic acid, ascorbic acid and GB´s. Has been reported these compounds can modulate the effect of frost stress by increasing the antioxidant capacity or compound as phenolics, reducing its oxidative damage and stabilizing cell tissues [1]. In fact, for new plantations of this species, exogenous application of compatible solute/osmolyte could potentially be one of the intervention strategies to improve plant freezing tolerance [2]. Thereby, GB plays an important role in cellular osmotic adjustment, which is associated with improved freezing tolerance through increase in resistance to freeze-induced dehydration/desiccation [3]. Also, it is known its role as a compatible solute that effectively stabilizes the quaternary structures of enzymes/proteins and maintain a highly ordered state of biological membranes and reducing cellular damage caused by freeze-thaw stress as reported in monocot species such as maize (Zea mays) [4] and wheat (Triticum aestivum) [5] showing successful reduction of oxidative stress and photoinhibition. The GB´s, synthesized at elevates rates in response to abiotic stresses, confer cryoprotective effects from its compatibility with macromolecular structure and function [6,7]. In addition, GB has received considerable attention as key osmoprotectant in mediating different plants responses, including growth, protein modification, photosynthesis, gene expression, and oxidative defense. Exogenous application of GB has shown promise for managing abiotic stress in plants, suggesting its involvement in protecting plant cells [6,7,8]. However, it should be note that in chestnut there are no reports regarding use of GB to increase the chilling tolerance (Figure 1B).
The use of osmoprotectant compounds such GB in young chestnut orchards from areas with a higher risk of frost during the first years of the training system formation, could be an alternative for agronomic management to prevent and mitigate severe damage to young tissues caused by low temperatures, particularly with a prolonged duration. In the last frost that occurred in January 2023 in full vegetative growth with a temperature of ‒2.34°C and a duration of 3.5 h, the damage caused by this event delayed the development of the trees for at least one season (Figure 2A,B). Because it is an extensive crop, the orchards do not have anti-frost system for frost control in critical phenological stages such as sprouting and new shoots growth. Therefore, it is necessary to looking forward for other lower-cost technologies such as these osmoprotectant compounds that allow preventing or mitigating the frost damage in critical phenological stages such as sprouting and growth of new shoots and leaves on new chestnuts orchards would become in a limitation of this crop developing in areas with a greater probability of frost occurrence.
2. Materials and Methods
The Centro de Fruticultura at the Universidad de La Frontera in Southern Chile has initiated a research line on fruit crop protection with osmoprotective compounds such as GB obtained from marine algae. The present study was the first to explore the hypothesis that the exogenous GB application on young chestnut plants that can mitigates freeze-thaw injury by reduction of oxidative stress as trees tolerance antioxidant mechanism.
2.1. Experimental Site and Treatments
The study was carried out during the 2022/23 season in an experimental chestnut three years-old trees (cultivar Cittá di Castello) orchard planted in Vilcún County (38° 41' 40" S; 72° 24' 57" W), La Araucanía region, Southern Chile. Orchard was conducted in a leader training system with planting distances between rows of 8 x 7m (178 trees ha‒1). The inter row were sowed with a perennial ryegrass (Lolium perenne) as cover crop to protect the soil and incorporate and recycle nutrients (Figure 1B). The treatments were the following: control (water; ‒GB) and GB application (+GB) at 120 g hL‒1 of commercial Intracell® (GB >96%) as osmoprotectant natural extract and sprayed in an output of 600 L ha-1 of water, determined by canopy volume (4πab2, a: ½ height; b: ½ width). Spraying was carried out using a back sprayer 17 L (Cifarelli 1200, Cifarelli S.p.A-Voghera, Italia) with an output of 5 L min‒1. Foliar application was carried out as preventive treatment by three times between mid and the end of spring prior to the occurrence of late spring and early summer frosts, common in the last three years, due to the climate variability of pre-Andes mountains where orchard was located. First preventive GB spraying was done in beginning of sprouting during the appearance of the leaves, phenological state highly susceptible to damage by low temperatures, below 0°C (spring, October). The second preventive application was carried out in leaf opening (spring, November), and third application was sprayed during the state of leaves free of stipules and meristematic apex in full development with herbaceous tissues susceptible to frost damage (early summer, december). It should be note that in the experimental chestnut orchard there was a frost for January 2022 and 2023 was ‒1.90 and ‒2.30°C, measured at a height of 1.5m with a duration of 3 and 2h respectively (Figure 2A). In fact, Figure 2B, shows for January the surface sensor (10cm from the ground) registered an absolute minimum temperature of ‒2.30°C for 2h, with an average relative air humidity above 90% during the time that the frost has occurred, which produced strong damage to the herbaceous tissues of chestnut trees without GB application.
2.2. Vegetative Growth
The height (m) and trunk diameter (mm) of trees were monthly measured in January and February, in full growth, and until March at the end of the growth stage. Measurement performed the same day for each month. For height, the trunk tagged at 20 cm from soil and determined by a measuring tape. Diameter was determined using a caliper (digital caliper CALDI-6MP, Truper, Mexico), similarly, to reported by Meriño-Gergichevich et al. [9]. For leaf dry matter, samples were weighed in an analytical balance (Model BA2204B, Biobase Meihua Trading, China) and disposed for drying at 70ºC in a forced-air oven for 48h (Heratherm OGS100, Thermo Scientific, Massachusetts, United States).
2.3. SPAD-Chlorophyll and Total Chlorophyll Content
For measurement of SPAD-chlorophyll contents (Chlorophyll Meter SPAD-502, Konica Minolta, Tokyo, Japan) six fully expanded leaves from tagged shoots were carefully measured. For the extraction and quantification of chlorophyll a, b and total chlorophyll the protocol of Wellburn [10] with minor modifications was used. Regarding total chlorophyll a+b, it was weighed 100 mg of leaves and 10ml of methanol (100%) was added for obtained chlorophyll extract leaf sample. Subsequently, it was homogenized in a vortex for 25s and then centrifuged at 6.000 rpm for 15 min at 4°C (Hermle; Z 300 K, Germany). Immediately, the supernatant was recovered and the process was repeated until all the color was extracted from the leaves sample. Subsequently, it was filtered (nylon 0.2 µm, 13mm-in diameter) to eliminate plant debris. Afterwards, the extract was collected in a 25mL volumetric flask, under dark conditions. Chlorophyll content was determined by absorbance at 653 and 666 nm, using a spectrophotometer (Jasco, V-730, Japan). The determination of chlorophyll a and b and total chlorophyll, was carried out according to the following:
Chl a (µg mL‒1) =15.65 A666-7.34 A653
Chl b (µg mL‒1) = 27.05 A653-11.21 A666
2.4. Net Photosynthesis, Stomatal Conductance and Transpiration
Photosynthesis (AN), stomatal conductance (gs) and transpiration rate (E) were registered with a portable Photosynthesis System Targas-1 (110 Haverhill Road, Suite 301 Amesbury, MA 01913 U.S.A), which contain compact, high precision, non-dispersive infrared gas analyzer for CO2 and H2O. The measurements were conducted between 1:00 and 3:00 PM, choosing a healthy leaf from the middle part of the tree, with an adequate size that allowed covering the space of the measuring clamp and exposed to sunlight. Subsequently, the middle of the leaf was introduced into the equipment´s clamp, until the measurement stabilized and yielded a net assimilation result on the equipment screen.
2.5. Total Phenols, Radical Scavenging Activity and Lipid Peroxidation
Total, phenolics concentration (TPC) was determined in the leaves following the method of Slimkard and Singleton [11] using the Folin-Ciocalteu reagent. Hundred fifty µL of ethanolic extracts (80% v/v) were mixed with 450 µL H2O and 100 µL of the Folin-Ciocalteu reagent. After 5 min, 300 µL of Na2CO3 (7% w/v) were added. The absorbance of the mixture was measured by a UV‒VIS spectrophotometer at 765 nm (UV‒VIS spectrophotometer SP 8001), Metertech Inc. Taipei, Taiwan) and TPC was expressed as mg of gallic acid equivalents (GAE) per g‒1 FW.
For radical scavenging activity (RSA) the 2,2– diphenil-1-picrylhydrazyl (DPPH) assay were used leaves samples (0.10g) were homogenized with 1mL of methanol (80% v/v) and centrifuged for 5 min at 10,000 rpm (4°C). The supernatant was collected and stored at ‒80°C until analysis. The free RSA of the methanol extracts was measured as absorbance decrease at 517 nm using Trolox as a standard and expressed as milligram of Trolox equivalent per gram of fresh weight (mg TEg‒1 FW)[12].
As oxidative stress indicator, the lipid peroxidation (LP) was determined in fresh leaves (0.15 g) using thiobarbituric acid reacting substances (Tbars) assay, according to the modified protocol of Du and Bramlage [13]. Absorbance was followed at 532, 600 and 440nm to correct the interference generated by Tbars-sugar complexes. The results were expressed as nanomol of equivalents of malondialdehyde (MDA) contents per fresh weight (nmol MDA g‒¹ FW), a secondary product of the polyunsaturated fatty acid oxidation.
2.6. Antioxidant Enzymes
For enzyme extracts, 0.6 g samples of leaf material were frozen in liquid nitrogen and then ground in 1.8 mL of 0.1 M potassium phosphate buffer (pH 7.0). Extracts were centrifuged (Lab. Companion, Korea) for 15 min at 13,000×g at 4 °C and the supernatant obtained was stored at ‒20°C until analysis. The assays were performed under UV light being the extracts previously treated with polyvinylpyrrolidone (PVPP) to remove polyphenols that could interfere with enzymatic determinations. Ten milligrams of PVPP were added to 400 μl of extract, shaken and centrifuged at 12,000xg for 10 min at 4°C, using the supernatant for enzymatic determinations of catalase (CAT), ascorbate peroxidase (APX), glutathione reductase (GR), peroxidase (POD) and superoxide dismutase (SOD) according to the method described by Aguilera et al. [14] and Carneiro-Carvalho et al. [15] with minor modifications. In addition, the supernatant obtained was used for protein quantification by Bradford [16]. A microplate spectrophotometer was used for all determinations (Synergy HTX UV‒visible Biotek, Winooski, VT, USA). Enzyme results were expressed as specific activity (nmol AU mg‒1 protein).
2.7. Experimental Design and Statistical Anayles
A randomized complete block design was employed with three replications (10 trees per replication). For dry matter, SPAD-Chlorophyll, total chlorophyll, TPC, RSA, LP and antioxidant enzymes data were statistically compared by a one-way analysis of variance (ANOVA). Whereas for tree height and diameter, net photosynthesis, stomatal conductance, and transpiration rate a two-way ANOVA was performed. A Tukey post-hoc test was used to determine the significance of differences among the means with a significance level at P≤0.05. All the statistical analyses were carried out with the free software R©, version 3.6.1.
3. Results and discussion
In relation to height (m)(Figure 3A), the highest values were observed for the +GB in the measurements carried out in January (28%), February (24%) and March (25%) compared to the ‒GB that achieved a lesser growth for the same evaluated time January (1.39m), February (1.52m) and March (1.65m)(P≤0.05). Various compatible solutes enable plants to tolerate environmental stress, and GB is one of the most studied among such solutes. It should be noted that on our studied chestnut plants +GB stimulated the vegetative growth of trees even after they have been suffered frost damage in summer. Glycine-Betaine is one of the most effective compatible solutes and it protects cellular structures from stresses by maintaining an osmotic balance and stabilizing the quaternary structures of complex proteins [18]. Concomitant with our findings, an increase in growth and development was observed in eggplant (Solanum melongena) [19] and in Strawberry (Fragaria x annanasa) cultivars Fortune and Albion treated with GB compared to control [8]. Likewise, a study carried out on Indian mustard (Brassica juncea), with foliar application of GB (20mM) increased the shoots length by 11.79% compared to the control [20]. From the published information, it is clearly seemed that GB modulates plant growth and development by phytohormones synthesis such as auxin, gibberellin, cytokinin, and salicylic acid. In different plant species under abiotic stress condition, such as wheat, corn, barley, tomato, as well as in model plants such as Arabidopsis, GB synthesized and accumulated and exogenously applied, increase the plant height, leaf area, shoot biomass, stem length, root length, flowering and fruit size [19, 20, 21, 22, 23, 24, 25, 26]. Regarding the trunk diameter (mm), there were no significant differences between ‒GB and +GB for measurements performed in January, February and March showing an average diameter of 29.06 mm by the end of summer (Figure 3B). Whereas the +GB leaves, showed the highest fresh (31.99g) and dry weight (12.78g) in in comparison to ‒GB leaves that reached a lower fresh and dry weight of 15.75 and 6.71g respectively. Moreover, dry matter (%), there were no significant differences between +GB (0.62) and –GB (0.60)(Figure 4). With respect the SPAD‒chlorophyll measurement (Figure 5A) neither were determined, no significant differences were found between the GB treatments (47.40 and 47.37 respectively). In relation to total chlorophyll there were no significant differences between +GB (2.76 mg g‒¹ FW) and ‒GB (2.43 mg g‒¹ FW)(Figure 5B). Chlorophyll is an important photosynthetic pigment, largely determining photosynthetic capacity and hence plant growth. Indeed, foliar application of GB in tomato, considerably enhanced chlorophyll content under stress condition [18, 23, 27]. The AN measured in the month of January was significantly higher for +GB (16.48 µmol CO2 m‒2 s‒1) treatment compared to the control 11.93 µmol CO2 m‒2 s‒1), respectively. However, the measurement carried out in February AN did not show significant differences between +GB (14.53 µmol CO2 m‒2 s‒1) and ‒GB (13.13 µmol CO2 m‒2 s‒1), respectively. Moreover, the measurement of AN carried out in March prior to leaf fall, presented significant differences between the treatment with GB and ‒GB (13.40 and 7.42 µmol CO2 m‒2 s‒1), respectively (Figure 6A). In a study conducted on mustard net photosynthetic rate was enhanced by +GB (20 mM)[20]. These results were concomitant with our measurements carried out in January and March (2023). Whereas, in rice GB treated seedlings, remained higher AN, possibly because of the protective effect of GB in different components of photosynthesis as well as phytohormones (auxin, brassinolide, gibberellin, and cytokinin) and molecules such as sucrose [28]. Moreover, sucrose accumulation has cryoprotective significance too and its elevation in GB treated plants may add to cold tolerance [29]. The highest gs was achieved in January with +GB (216 mmol H2O m-2 s-1) exhibiting a 44% higher gs than ‒GB (P≤0.05), whereas for February and March gs in +GB was increased a 39 and 33% respectively compared to ‒GB (P≤0.05)(Figure 6B). Our study showed similar results with those obtained by Hamani et al. [26] in cotton (Gossypium sp.), who reported an increased gs with +GB (5mM) in plants under salinity stress. Exogenous applications of GB in cape gooseberry (Physalis peruviana L) at a dose of 100mM showed higher gs (95 mmol H2O m‒2 s‒1) values regarding the control under waterlogging condition [30]. A study carried out in strawberry demonstrated that exogenous application of antifreeze protein (AFP) type III and GB (100mg L‒1) increased gs from 179.9 to 619.4 mmol H2O m‒2s‒1 for Sweet Ann and Karbala cultivars. Results of the study showed that AFP and GB applications may have contributed through diverse ways to tolerance to low temperature and these compounds are potentially useful for inducing tolerance to cold temperature in studied plants [8]. January is the warmer month in summer for southern Chile concomitantly with the highest E value was obtained for the +GB treatment (3.51 mmol H2O m‒2 s‒1) a 32% higher than determined for ‒GB (2.39 mmol H2O m‒2 s‒1)(P≤0.05)(Figure 6C). Similarly, February also showed in +GB a significant (27%) higher E in comparison to –GB, whereas in March the leaves of chesnut showed similar E values in both GB treatment averaging 2.71 and 2.06 mmol H2O m‒2 s‒1 respectively. The result of our work agrees with a study carried out in cotton (Gossypium hirsutum), in which they obtained an increase in transpiration rate through the foliar application of GB (5mM) [26].
With respect to antioxidant determination, ‒GB leaves harvested during the last measurement (March) showed TPC significantly higher content (35%) in relation to +GB leaves (60.96 and 39.23 mg GAE g‒1 FW)(P≤0.05; Figure 7A). Contrarily, a study carried out in lettuce (Lactuca sativa) with exogenous application of GB (25mM) enhanced the total phenols compared to the ‒GB treated plants [31]. Moreover, high levels of TPC were observed in GB (20 mM) treated pomegranate (Punica granatum) fruits during cold storage in postharvest, as effective alleviation of chilling injury [32]. Similarly to reported by Molei et al. [32], +GB presented a 37% higher DPPH activity than determined for ‒GB (33.50 mg TE g‒1 FW)(P≤0.05; Figure 7B). Whereas our study demonstrated that higher RSA, involving non enzymatic and enzymatic antioxidant should be related in amelioration of oxidative stress.
In relation to antioxidant enzymatic performance determined in chestnut leaves, our experiment showed that CAT, APX and POD were 95, 50 and 44% higher in –GB than in +GB treated trees respectively (P≤0.05; Figure 8A,B,D). Whereas GR was 65% increased by +GB in comparison to those –GB treated leaves (P≤0.05; Figure 8C), but SOD was not modified showing activities from 253±26 to 296±18 U min‒1 FW (Figure 8E). Enzymatic antioxidants are activated against oxygen radical triggering oxidative stress when vegetables are subjected to environmental stresses. However, in our study SOD was similar both in leaves collected from –GB and +GB trees, although this enzyme represents the first line protecting against superoxide radical by dismutation to hydrogen peroxide (H2O2). A study carried out on flag leaves of winter wheat, +GB application showed significant increases in the activity of antioxidant enzymes SOD, CAT and POD under limited irrigation conditions, highlighting the role of GB on membrane stabilization and modulation amounts of H2O2 [33]. Whereas subirrigation with GB (30mM) on cabbage (Brassica oleraceae L) 14‒days‒old seedlings under stressor conditions due to freezing temperatures determined a greater antioxidant activity of the plants with a higher activity of antioxidant enzymes CAT, APX, and SOD and glutathione, as non-enzymatic antioxidant [34]. In maize (Zea may), soybean (Glycine max) the application of exogenous GB was the most effective among all compounds to enhance the drought tolerance with reduced levels of reactive oxygen species (ROS), increased activities of antioxidant enzymes and higher accumulation of osmolyte contents [35,36]. It should be noted that GB may be involved in the inhibition of ROS accumulation, protection of photosynthetic machinery, accumulation of compatible solutes to maintain turgidity in cells, activation of some stress related genes, and protection of cell membrane and quaternary structure of proteins [37,38]. All these studies have reported that GB participate as an osmolyte, enhancing resistance against osmotic stress through increase in the osmolality of the cell [34,35,36].
In relation to the lipid peroxidation, the highest content was presented for –GB treatment (90.03 nmol MDA g‒1 FW) with significant differences with respect to the +GB application, that showed a 42% lower LP in leaves (51.9 nmol MDA g‒1 FW)(Figure 8C). It should be noted that our work agrees with a study carried out in pomegranate fruits, cv. Malese Saveh during cold storage in which it was determined that the application of exogenous GB (20mM) achieved a lower content of MDA compared to the control (untreated) [31]. As similarly to study carried out on wheat, showed that exogenous application of GB significantly reduced the accumulation of MDA [33] and a study carried out in maize the exogenous application of osmoprotective compounds decreased substantially the ROS and MDA content [35]. Furthermore the exogenous application of GB in cabbage showed a lower content of MDA after environmental stress due to low temperatures (from ‒2.5 to ‒3.5°C)[34]. The measurement of MDA content has long been used as a lipid peroxidation marker in studies related to oxidative stress and redox signaling, particularly in those studies focused on plant responses to abiotic stresses [38,39,40,41]. Lower levels of MDA could be well associated with lesser cell oxidation in chesnut tissues under stressor climatic variability.
4. Conclusions
Osmoprotective compounds, such as Glycine-Betaine has become as an interesting agronomic tool to increase tolerance to environmental stresses such as low temperatures that cause damage to herbaceous shoots and leaves in young chestnut trees. However, these results are preliminary and therefore, it is necessary to continue with the studies to refine the doses, number of foliar and sub-irrigation applications before and after the occurrence of climatic adversities on critical phenological stages such as sprouting and growth of new shoots and leaves, especially in new orchards located in Andean foothill territories of Southern Chile.
Conflicts of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationship as a potential conflict of interest.
Author Contributions statement
ME, AG, CM-G and IR designed and supervised the study. CM-G, LM, GG-C and AR analyzed the data. CM-G, ME, AG, IR, LM and GG-C wrote the manuscript, and all authors critically revised the manuscript and approved the final version.
Acknowledgments
This study was supported by the Regional Government of La Araucanía (GORE), Universidad de La Frontera DIUFRO DI22-45 and DIUFRO DI22-2001. We thanks to technical support from the Laboratory of Plant Physiology and Nutrition in Fruit Crops at the Universidad de La Frontera. Special thanks to Dr. Anita Sandoval from INIA for her collaboration in fine-tuning the protocol for chlorophylls and technicians Francisca Cabrera and Guillermo Contreras from INIA for the collaboration in the agronomic management of the chesnut orchard.
References
- Jalili, I.; Ebadi, A.; Ashari, M.; Kalatehjari, S.; Ali-Aazami, M. Foliar application of putrescine, salicylic acid and ascorbic acid mitigates frost stress damage in Vitis vinifera c.v ´Giziluzum. BMC Plant Biol. 2023, 23, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Arora, R. Mechanism of freeze‒thaw injury and recovery: A cool retrospective and warming up to new ideas. Plant. Sci. 2018, 270, 301–313. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarnillo, F.; Shigematsu, S.; Minami, A.; Kawamura, Y. Spatial and temporal profile of glycine‒betaine accumulations in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.P.; Li, P.H.; Chen, T.H. Glycine betaine increases chilling tolerance and reduces chilling‒induced lipid peroxidation in Zea mays L. Plant Cell Environ. 2000, 23, 609–618. [Google Scholar] [CrossRef]
- Allard, F.; Houde, M.; Kröl, M. Betaine improves freezing tolerance in wheat. Plant Cell Physiol. 1998, 39, 1194–1202. [Google Scholar] [CrossRef]
- Koster, K.L.; Lynch, D.V. Solute accumulation and compartmentation during the cold acclimation of Puma rye. Plant Physiol. 1992, 88, 108–113. [Google Scholar] [CrossRef]
- Zulfiqr, F.; Ashraf, M.; Siddique, K.H. Role of glycine betaine in the thermotolerance of plants. Agronomy 2022, 12, 276. [Google Scholar] [CrossRef]
- Aras, S.; Eşitken, A. Effects of antifreeze proteins and glycine betaine on strawberry plants for resistance to cold temperature. IPCBEE 2013, 60, 21. [Google Scholar]
- Meriño-Gergichevich, C.; Luengo-Escobar, A.; Alarcón, D.; Reyes-Díaz, M.; Ondrasek, G.; Morina, F.; Ogass, K. Combined spraying of boron and zinc during fruit set and premature stage improves yield and fruit quality of European hazelnut cv. Tonda di Giffoni. Frontiers in Plant Science 2021, 12, 984. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorohyll a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Slimkard, K.; Singleton, V.L. Total, phenol analysis: Automation and comparison with manual methods. Am. J Enol. Vitic. 1997, 28, 29–55. [Google Scholar]
- Yu, L.; Haley, S.; Perret, J.; Harris, M.; Wilson, J.; Qian, M. Free radical scavening propierties of wheat extracts. J. Agric. Food Chem. 2002, 50, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1556–1570. [Google Scholar] [CrossRef]
- Aguilera, A.; Tereucán, G.; Ercoli, S.; Cornejo, P.; Gomez, M.R.; Uhlmann, L.; Ruiz, A. Influence of organic and chemical fertilisation on antioxidant compounds profiles and activities in fruits of Fragaria ananassa var. Camarosa. J. Soil Sci. Plant Nutr. 2020, 20, 715–724. [Google Scholar] [CrossRef]
- Carneiro-Carvalho, A.; Aires, A.; Anjos, R.; Martins, L.; Pinto, T.; Peixoto, F.; Gomes-Laranjo, J. The role of silicon fertilization in the synthesis of phenolic compounds on chestnut plants infected with P. cinnamomi and C. parasitica. J. Plant Dis. Protect. 2020, 127, 211–227. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Murata, N. Glycinebetaine protects plants against stress: Mechanism and biotechnological application. Plant. Cell. Environ. 2011, 34, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.; Zhang, J.; Jing, L.; Xiaping, G.; Ma, H.; Yanqiang, G.; Chang, Y.; Xie, J. Effects of exogenous glycine betaine and cycloleucine on photosynthetic capacity, amino acid composition, and hormone metabolism in Solanum melongena L. Sci. Rep. 2023, 13, 7626. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.; Parrey, Z.A.; Shah, S.H.; Mohammed, F. Glycine betaine mediated changes in growth, photosynthetic efficiency, antioxidant system, yield and quality of mustard. Scientia Hortic. 2021, 285, 110170. [Google Scholar] [CrossRef]
- Yang, X.; Liang, Z.; Lu, C. Genetic engineering of the biosynthesis of glycinebetaine enhances photosynthesis against high temperature stress in transgenic tobacco plants. Plant Physiol. 2005, 138, 2299–2309. [Google Scholar] [CrossRef]
- He, C.; Zhang, W.; Gao, Q.; Fang, A. Enhancement of drought resistance and biomass by increasing the amount of glycine betaine in wheat seedlings. Euphytia 2011, 177, 151–167. [Google Scholar] [CrossRef]
- Che-um, S.; Samphumphiang, T.; Kirddmanee, C. Glycinebetaine alleviates water defficit stress in indica rice using proline accumulation, photosynthetic efficiencies, growth performances and yield attributes. Aust. J. Crop Sci. 2013, 7, 213–218. [Google Scholar]
- Ke, Q.; Wang, Z.; Ji, C.Y.; Jeong, J.C. Transgenic poplar expressing codA exhibits enhanced growth and abiotic stress tolerance. Plant Physiol Biochem. 2016, 80, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Manaf, H.H. Benefical effects of exogenous selenium, glycine betaine and seawed extract on salt stressed cowpea plant. Ann Agric Sci. 2016, 61, 41–48. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Li, S.; Chen, J.; Ami, A.S.; Wang, S.; Xiaojun, S.; Zain, M.; Gao, Y. Linking exogenous foliar application of glycine betaine and stomatal characteristics with salinity stress tolerance in cotton (Gossypium hirsutum L.). seedlings. BMC Plant Biol. 2021, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Zhang, W.; Wang, C. Genetic engineering of the biosyntesis of glycinebetaine leads to alleviate salt-induced potassium efflux and enhances salt tolerance in tomato plants. Plant Sci. 2017, 257, 74–83. [Google Scholar] [CrossRef]
- Zhang, T.; Liang, J.; Wang, M. Genetic engineering of the biosynthesis of glycinebetaine enhances the fruit development and size of tomato. Plant Sci. 2019, 280, 355–366. [Google Scholar] [CrossRef]
- Syeed, S.; Sehar, Z.; Masood, A.; Anjum, N.A.; Khan, N.A. Control of elevated ion accumulation, oxidative stress, and lipid peroxidation with salycilic acid-induced accumulation of glycine betaine in salinity-exposed Vigna radiata L. Apply Biochem Biotechnol. 2021, 193, 3301–3320. [Google Scholar] [CrossRef]
- Castro-Duque, N.E.; Chávez-Arias, C.C.; Restrepo-Díaz, H. Foliar glycine betaine or hydrogen peroxide sprays ameliorate waterlogging stress in cape gooseberry. Plants 2020, 9, 644. [Google Scholar] [CrossRef]
- Shams, M.; Yildirim, E.; Ekini, M.; Turan, M.; Dursun, A.; Parlakova, F.; Kul, R. Exogenously applied glycine betaine regulates some chemical characteristics and antioxidative defence systems in lettuce under salt stress. Hort. Environ. Biotech. 2016, 57, 225–231. [Google Scholar] [CrossRef]
- Molei, S.; Rabiei, V.; Soleinami, A.; Razave, F. Exogenous application of glycine betaine increases the chilling tolerance of pomegranate fruits cv. Malese Saveh during cold storage. J. Food Process Preserv. 2021, 45, e15315. [Google Scholar]
- Khedr, R.A.; Sorour, S.G.R.; Aboukhadrah, S.H.; El Shafey, N.M.; Abd Elsalam, H.E.; El-Sharnouby, M.E.; El-Tahan, M. Alleviation of salinity stress effects on agro-physiological traits of wheat by auxin, glycine betaine, and soil additives. Saudi J. Biol Sci. 2022, 29, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Zhang, Y.; Li, K.; Zhou, Y.; Zhang, M.; Li, Z. Exogenous application of glycine betaine improved water use efficiency in winter wheat (Triticum aestivum L.) via modulating photosynthetic efficiency and antioxidative capacity under conventional and limited irrigation conditions. Crop J. 2019, 7, 635–650. [Google Scholar] [CrossRef]
- Min, K.; Cho, Y.; Kim, E.; Lee, M.; Lee, S.R. Exogenous glycine betaine application improves freezing tolerance of cabbage (Brassica oleracea L.) leaves. Plants 2021, 10, 2821. [Google Scholar] [CrossRef] [PubMed]
- Shemi, R.; Wang, R.; Geith, E.M.S.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of salicylic acid, zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress. Scientific Reports 2021, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Andrade, C.A.; de Souza, K.R.D.; de Olivera Santos, M.; da Silva, D.M.; Alves, J.D. Hydrogen peroxide promotes the tolerance of soybean to waterlogging. Sci. Hortic. 2018, 232, 40–45. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Shomo, M.; Tobita, S. Effects of proline and betaine in heat inactivation of ribulose-1, 5-bisphosphate carboxylase/oxigenase in crude extract of rice seedlings. Photosynthetica 2000, 36, 557–563. [Google Scholar] [CrossRef]
- Palonem, P.; Buszard, D.; Donnelly, D. Changes in carbohydrates and freezing tolerance during cold acclimation of red raspsberry cultivars grown in vitro and in vivo. Physiol. Plant 2002, 110, 393–401. [Google Scholar] [CrossRef]
- Giri, J. Glycine betaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1745–1751. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the bioma scale. Front. Ecol. Evol 2018, 6, 64. [Google Scholar] [CrossRef]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Evaluated chesnut tree with frost damage (A) and without damage (B). Cultivar Cittá di Castello planted at Vilcún (38°41'40"S; 72°24'57"W), La Araucanía region, Southern Chile.
Figure 1.
Evaluated chesnut tree with frost damage (A) and without damage (B). Cultivar Cittá di Castello planted at Vilcún (38°41'40"S; 72°24'57"W), La Araucanía region, Southern Chile.

Figure 2.
(A) Monthly temperatures (°C) and frost number, mean frost duration (h) and (B) frost summatory (<0.0°C h month‒1) in chesnut orchard cultivar Cittá di Castello planted at Vilcún (38°41'40"S; 72°24'57"W), La Araucanía region, Southern Chile.
Figure 2.
(A) Monthly temperatures (°C) and frost number, mean frost duration (h) and (B) frost summatory (<0.0°C h month‒1) in chesnut orchard cultivar Cittá di Castello planted at Vilcún (38°41'40"S; 72°24'57"W), La Araucanía region, Southern Chile.

Figure 3.
Monthly measurement of (A) tree height (m) and (B) trunk diameter (mm) in chesnut sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05) between treatments for the same measurement time.
Figure 3.
Monthly measurement of (A) tree height (m) and (B) trunk diameter (mm) in chesnut sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05) between treatments for the same measurement time.

Figure 4.
(A) Leaf dry weight (g kg‒1 DW) and (B) Dry matter (%) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
Figure 4.
(A) Leaf dry weight (g kg‒1 DW) and (B) Dry matter (%) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
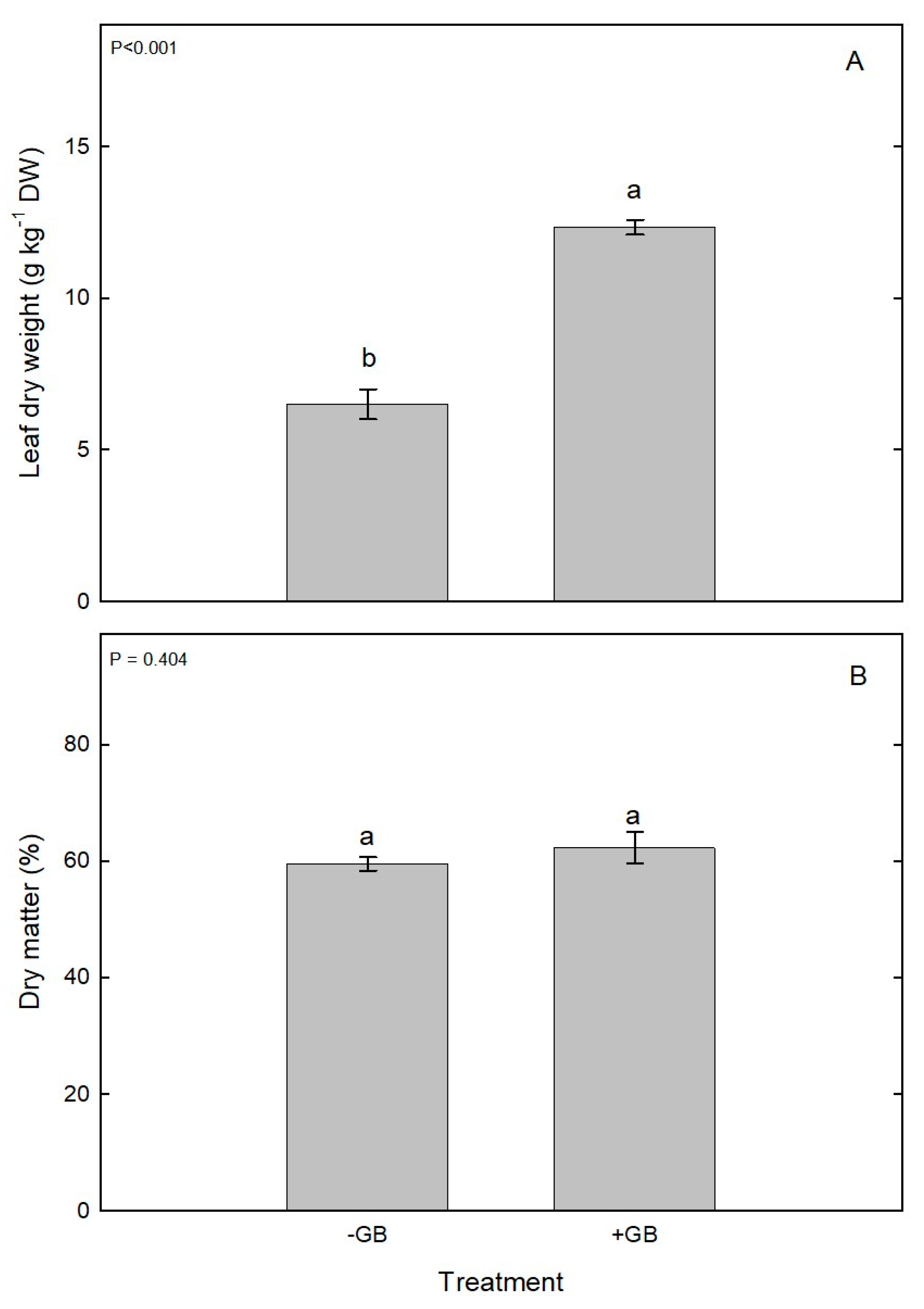
Figure 5.
(A) SPAD-chlorophyll and (B) Chlorophyll content in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
Figure 5.
(A) SPAD-chlorophyll and (B) Chlorophyll content in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
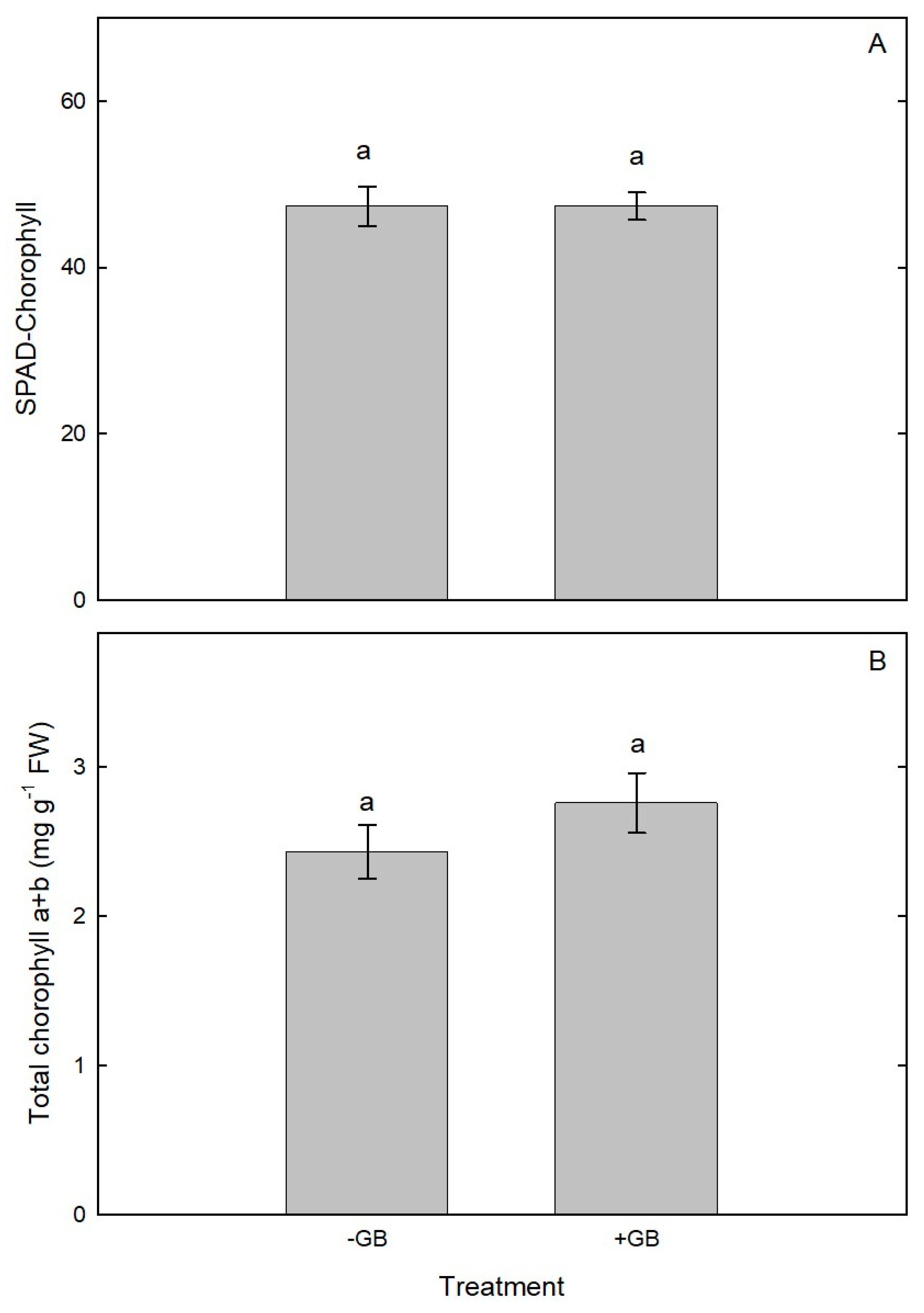
Figure 6.
(A) Photosynthetic activity (µmol CO2 m‒2 s‒1), (B) stomatal conductance (mmol H2O m‒2 s‒1) and (C) transpiration rate (mmol H2O m‒2 s‒1) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05) between treatments for the same measurement time.
Figure 6.
(A) Photosynthetic activity (µmol CO2 m‒2 s‒1), (B) stomatal conductance (mmol H2O m‒2 s‒1) and (C) transpiration rate (mmol H2O m‒2 s‒1) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05) between treatments for the same measurement time.

Figure 7.
(A) Total phenol content (mg GAE g‒1 FW), (B) Radial scavenging activity (mg TEg‒1 FW) and Lipid peroxidation (nmol MDA g‒¹ FW) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
Figure 7.
(A) Total phenol content (mg GAE g‒1 FW), (B) Radial scavenging activity (mg TEg‒1 FW) and Lipid peroxidation (nmol MDA g‒¹ FW) in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).

Figure 8.
Activity of (A) CAT, (B) APX, (C) GR (D) POD (nmol min‒1 mg DW) and (E) SOD (U min‒1 FW) antioxidant enzymes in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).
Figure 8.
Activity of (A) CAT, (B) APX, (C) GR (D) POD (nmol min‒1 mg DW) and (E) SOD (U min‒1 FW) antioxidant enzymes in leaves of chesnut trees sprayed with GB during spring (2022). Cultivar Cittá di Castello planted at Vilcún, La Araucanía region, Southern Chile. Bars represent mean of three replications ± S.E. Different lowercase letters indicate statistically significant differences (P≤0.05).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.


Melatonin and Sodium Nitroprusside Impact on Vegetative Growth, Physiological, Biochemical Traits and Mineral Elements Contents of California Wonder Green Bell Pepper (Capsicum annuum) Under Different Temperatures
Dariush Ramezan
et al.
,
2023



MDPI Initiatives
Important Links
© 2024 MDPI (Basel, Switzerland) unless otherwise stated