
Preprint
Article
Assessment of the divergent influence of natural and non-seasonal hydrological fluctuations on functional traits and niche characteristics of plant guilds along the Xiangxi River, China
Altmetrics
Downloads
74
Views
19
Comments
0
A peer-reviewed article of this preprint also exists.




This version is not peer-reviewed
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Alerts
Abstract
The reservoir water level fluctuation zones (RWLFZs) and the natural riparian zones (NRZs) are two riparian ecosystems with the dramatically opposite hydrological rhythms which notably influence the plant guilds. However, little is known about the discrepancy of functional traits and niche characteristics of plant guilds in the RWLFZ and NRZ under different flooding rhythms. Here the RWLFZs (degraded riparian zones) and NRZs were chosen in the Xiangxi River Basin of the Three Gorges Reservoir (TGR) to investigate the variation in plant community structure, the spatial patterns of diversity, key functional traits and niche characteristics of dominant plant populations. The results showed that a total of 78 vascular plant species were identified in the riparian zones of Xiangxi River Basin. Most of the dominant species were annual herbs in the two riparian ecosystems and the percentage of annual dominant species increased from 65.79% in the NRZ to 67.34% in the RWLFZ. Compared with the NRZ, the specific leaf area, vegetation coverage and the aboveground biomass in the RWLFZ significantly increased by 74%, 27% and 19.6% respectively, while the water use efficiency of the RWLFZ decreased by 59.6% and there was no significant difference in the net photosynthetic rate between them, which showed that annuals in the RWLFZ adopt the R adaptation strategy with fast-growing, short lifespan and quickly acquiring and investing resources by altering leaf morphology such as expanding leaf area. The Simpson dominance index of RWLFZ was significantly higher than that of NRZ. Thus counter-seasonally hydrological alterations had the significant effects on functional traits of dominant species in the RWFLZ. Moreover, the highly adaptable and widely distributed species with larger niche breadths and high important values usually had a higher niche overlap value in the RWLFZ than in the NRZ, which showed that the competition in the plant community after experiencing anti-seasonal flooding was much more intraspecific than interspecific and would facilitate the expansion of species niches. And the distribution of the plant community was mainly driven by elevation (flooding depth), flooding duration and TK in the WLFZs. Our findings imply that the species with large niche breadths and high important values should be prioritized in ecological restoration efforts in the newly formed hydro-fluctuation zone of the TGR.
Keywords:
Subject: Environmental and Earth Sciences - Ecology
1. Introduction
The operation of reservoirs exerts a substantial influence on the plant community, such as species composition, community structure and distribution patterns [1,2]. It is estimated that two-thirds of the world’s fresh water flowing to the oceans is obstructed by more than about 50,000 large dams—defined as those higher than 15 m in operation worldwide [3]. Ali demonstrated that the plant species in the water level fluctuation zones (WLFZs) of the Nubia Reservoir in Sudan, Africa, decreased sharply with only half of the plant species existed after 30 years of water storage [4]. Moreover, dams anticipate large-scale plant population disappearance and loss of terrestrial and arboreal species in submerged lowland forest areas [1]. So far, however, few experimental studies have been conducted to test the effect of water level fluctuations on the functional traits and niche characteristics of dominant species of plant communities in the reservoir water level fluctuation zones (RWLFZs) with a large vertical height difference compared with the natural riparian zones (NRZs). Therefore, it is of great significance to study the functional traits and niche of dominant species for evaluating the effects of the reservoir impoundments on vegetation community characteristics, for better prediction of RWLFZ plant community dynamics and for restoration efforts of riparian ecosystems.
The Three Gorges Dam (TGD), the world’s largest hydroelectric project, was built on the Yangtze River in China. Three Gorges Dam (TGD) began operating in June 2003, and the water level of the Three Gorges Reservoir (TGR) rose from approximately 60 m above sea level (asl) to 135 m asl above the original riverbank. The second impoundment occurred in October 2006, and the water level rose to 156 m asl. With the full operation of the TGD in 2010, the water level fluctuates from 145 m asl. in summer (May to September) to 175 m asl in winter (October to April) with a vertical height difference of 30 m in the reservoir, resulting in formation of an unnatural RWLFZ with a total area of 350 km2 [5]. The reversal of flooding time and prolonged submergence duration have dramatically altered the hydrological regimes and plant community in the RWLFZs [5,6,7]. For example, before the construction of Three Gorges Dam (TGD),the vertical drop of the NRZs of the main stream of Yangtze River was 10 ~ 47 m asl. and the NRZ was only inundated by natural summer flooding [8,9,10]. Thus, the complete operation of TGD generated different types of WLFZs, including the RWLFZs and the NRZs. For the RWLFZ, it had not experienced flooding before the impoundment of the TGR, and the vegetation in the RWLFZ is subjected to the anti-seasonally hydrological rhythm (i.e., summer exposure and winter inundation). The newly-formed ecosystems in the RWLFZs offer special challenges to restoration managers because of the reversal of flooding time and prolonged submergence duration, their short growing seasons, and long recovery periods.
Hydrological regime alterations anticipate a series of unfavorable vegetation changes, such as vegetation type, distribution patterns, community species composition and diversity, coverage and functional traits of plant guilds [11,12]. Riverine plant communities exhibit varied responses to diverse water level regimes [13]. The impoundment at the TGR is likely to exert distinct influences on the various vegetation types within the WLFZs. Furthermore, the duration of flooding is identified as a significant factor impacting species distribution. [14]. After 20 times of flood-dry cycles in the RWLFZ of the TGR since the impoundment in 2003, some previous dominant species that had not experienced multiple impoundments gradually died out, showing a certain degree of degradation, and some new dominant species would appear and grow more and more vigorously [13]. These alterations would accelerate the disappearance of original vegetation, especially trees and shrub, the formation of permanent waters and trigger the appearance of novel plant communities which gradually adapt to the environment of the RWLFZ in the TGR [12,15,16]. The species composition and structure of the plant community is still extremely unstable [17]. Zhang et al.[18] documented that perennial plants in the reservoir's riparian zone exhibited a markedly higher photosynthetic capacity per unit area, leaf nitrogen concentration, and stem mass ratio compared to their upland counterparts. However, little is known about the effects of prolonged submergence (4–8 months) and the reversal of inundation time of TGR on functional traits and niche characteristics of plant guilds in different WLFZs.
Previous investigations have noted that specific plant species are capable of withstanding extended periods of winter flooding in the Three Gorges Reservoir Area, facilitated by a "physiological compensation" mechanism that enhances their photosynthetic capacity [18,19]. Perennial vegetation within the WLFZ displayed a notably enhanced area-based photosynthetic capacity when compared to upland species. In addition, these WLFZ perennials exhibited significantly greater stomatal conductance, leaf nitrogen concentration, and stem mass ratio than their upland counterparts [18]. However, it remains largely unclear whether dominant plant species in the RWLFZs altered functional traits in response to the flood-dry-flood alternate changes after 20 cycles of operation of the TGR compared with the NRZs.
Niche theory has become an important means of evaluating biodiversity mechanisms, species coexistence and competition and the status of populations in natural plant communities [16,20]. The niche may be well characterized by two important niche indices, i.e., niche breadth and niche overlap [16,21]. The size of niche breadth of species depends on the utilization to environmental resources and adaptive capacity of species to environments [22]. The changes of soil environments caused by anthropogenic activities would lead to the competition for limited resources among plant species and their good adaptability to environments [23]. Because niche breadth and overlap can be quantitatively evaluated, they can effectively reflect the range of resource characteristics across which a species exists in its specific ecosystem, and indicate the extent that species utilize different types of resources and measure potential competition between species in natural plant communities [12,16]. The optimal fundamental niche of a species can enhance our comprehension of the underlying mechanisms that drive variations in abundance patterns, dominance, coexistence among plant species, and biodiversity across flood gradients. [24]. In the present study, two types of the WLFZs (i.e. RWLFZ and NRZ) with different hydrological regimes in the Xiangxi River were selected to investigate the changes in plant community structure, functional traits, niche characteristics of dominant plant populations and try to address the following specific questions:
- (1)
- What are the divergent influence of counter-seasonally and naturally hydrological regimes on the plant community species composition and functional traits in the two types of the WLFZs (i.e., RWLFZ and NRZ) in the Xiangxi River of the TGR?
- (2)
- What are the divergent niche characteristics of dominant species of plant communities in the two types of the WLFZs (i.e. RWLFZ and NRZ) and the mechanisms of competition of dominant plant guilds in the Xiangxi River of the TGR?
- (3)
- What are the ecological adaptation strategies of plant guilds based on functional traits and niche characteristics in the different types of the WLFZs of the TGR?
This work will contribute significantly to identifying the most suitable plant species for ecological restoration and re-vegetation endeavors in the unique riparian forests of the TGR and analogously environmentally delicate ecosystems.
Study area and research method
2.1. Study area overview
The Xiangxi River is located in the western part of Hubei Province (with a longitude from 110°25'E to 111°06'E and a latitude from 31°04'N to 31°34'N)[73], originating from the Shennongjia Forest District, with a total length of 97.3 km. The river mouth is 34.5 km upstream from the TGD, and the basin area is 3183 km2 with a natural drop of 1054 m[71]. Since the impoundment of the TGD, the Xiangxi River has transformed from a natural river channel into a reservoir-type river. When the TGR reaches a water level of 175 m, the bay's backwater length is approximately 42 km[72].The region has a subtropical continental monsoon climate, with distinct seasons and abundant rainfall. Precipitation within the basin decreases from northwest to southeast, with an annual rainfall ranging from 1440 to 1000 mm. The rainy season is mostly concentrated between June and September. The main types of soil are yellow brown soil and lime soil. The undulating topography and high hills account for more than eighty percent of the total area [25]. Before submergence, vegetation in the riparian forests was dominated by annuals, i.e., Setaria viridis, Digitaria ciliaris, and Leptochloa chinensis, perennials including Cynodon dactylon, Hemarthria altissima, and Capillipedium assimile, and trees such as Ficus tikoua, Pterocarya stenoptera, and Vitex negundo [6]. However, after the prolonged submergence, the number of plant species is strongly reduced and annual plants, i.e., Echinochloa crusgalli and Bidens tripartita, and perennials including C. dactylon are dominant species [6].
2.2. Field investigation and sampling
Vegetation surveys were conducted across nine transects from June to September in 2023 (Figure 1). Of these, there are six transects in the RWLFZs and three in the NRZs. The surveys were carried out during the flood-regulation periods, with each transect spanning widths between 40 to 60 m.. The water level fluctuated between 145 m and 175 m. Therefore, we divided each transect into three elevation zones (145-155 m, flooding depth 20-30m; 155-165 m, flooding depth 10-20m and 165-175 m, flooding depth 0-10m) in the RWLFZs and one elevation zone (175-180 m, no flooding) in the NRZs. On elevation zone of each transect, five 1×1m square sampling plots were established with a 20 m spacing between each plot. Soil density ( 0-20cm ) was measured by the cutting ring method. At each transect's elevation zone, three topsoil samples from 0-20 cm depth were randomly gathered and homogeneously combined to create a single composite sample for analysis... A total of 105 quadrats were sampled and five samples were mixed in each quadrat, sealed in plastic bags and brought to the laboratory. The name, number, height, coverage, frequency and above-ground biomass of each plant species in each quadrat were recorded, and finally harvested all plants to measure the above ground biomass of vegetation in each quadrat. The sketch map of study area and sampling sections are shown in Figure 1, and the basic characteristics of the quadrats is shown in Table 1.
2.3. Leaf functional traits
The Li-6400 portable photosynthetic equipment (Li-cor, Inc, USA) was utilized to measure photosynthesis parameters in the field at 9:00–11:00 or 15:00–17:00 on a clear day to avoid midday depression of photosynthesis. Following the measurement, the measured blades' markers (1-9) were collected and stored in an envelope bag (the envelope bag was labeled with sampling point information), and the labels were sorted and mailed back to the laboratory for measurement of further indicators. Photosynthesis parameters comprise the net photosynthesis rate (Pn), transpiration rate (Tr), stomatal conductance (Cs), inter-cellular CO2 concentration(Ci), leaf temperature (Tl), atmosphere temperature (Ta), atmosphere CO2 concentration (Ca), water use efficiency (WUE, Pn/Tr), etc. The unfolded leaves were collected in the laboratory in time, scanned, and saved by Canon Scan Li DE 110 scanner, and the leaf area of each leaf was calculated by Image J software (cm2), and the blade thickness was measured by avoiding the main vein with a spiral micrometer [26]. The scanned leaves were placed in the oven and dried to constant weight at 75℃. The dry matter weight of each leaf (g) was weighed and recorded (Precision: 0.001 g). The specific leaf area ((leaf area/leaf dry weight, cm2/g) of each plant species in various quadrats was calculated and recorded [27].
2.4. Environmental factors
Soil samples were prepared for analysis by air-drying and passing them through a sieve with a mesh size of less than 2 mm.. The pH value of soil was determined by potentiometric method [28].The determination of soil organic matter(OM)content is conducted using the dichromate volumetric method [29]. Total phosphorus (TP) and Available phosphorus (AP) were measured by molybdenum-antimony anti-spectrophotometric method [64]. Total phosphorus(TK) and Available potassium (AK) were determined by flame photometric method [30]. Ammonium(NH4+-N) and Nitrate(NO3--N) were determined using spectrophotometry. [31]. The soil moisture (SM) was measured using the oven-drying method,The heterogeneity of soil environmental factors in different elevations in the TGR were shown in Figure 2
2.5. Data analysis
2.5.1. Importance value
Within each vegetation assessment, individual samples were categorized to represent distinct resource states. The importance value (Pi) for each species was derived from a combination of their relative coverage (RC), relative frequency (RF), relative height (RH), and relative density (RD) across various quadrats. These metrics were then utilized as parameters to identify the predominant herbaceous species within the water level fluctuation zones and to conduct ecological niche analysis [32]. The computation is articulated by the following formula:
Where Pi is the importance value of the i-th species, RC is calculated as the projective coverage of a species divided by the sum of coverages of all species across all quadrats. RF is the count of occurrences of a species divided by the total occurrences of all species in all quadrats. RH is the average height of a species divided by the total height of all species across all quadrats. RD is the total count of individuals of a species divided by the total individual count of all species in all quadrats.
2.5.2. Species diversity index
Community alpha diversity was assessed through a suite of indices, including species richness (S), the Shannon diversity index (H), Pielou's evenness index (E), and Simpson's dominance index (D). Shannon Diversity Index represents the species diversity of the community, including species richness (number of species ) and evenness (distribution uniformity of individual number of each species). Pielou 's evenness index is used to measure the uniformity of individual number distribution of each species in the community. The value of this index is between 0 and 1. The higher the value is, the more uniform the distribution of each species in the community is. The Simpson 's dominance index reflects the uneven distribution of species or the concentration of dominant species in the community. The value of the index is between 0 and 1. The smaller the value, the more uniform the distribution of species in the community, and there is no obvious dominant species. The respective calculation formulas are presented below [33]:
Shannon diversity index:
Pielou evenness index:
Simpson dominance index:
Where Pi is the importance value of the i-th species, S represents the number of distinct species that are found within the quadrat, and N signifies the total number of species that are present within the quadrat
2.5.3. Niche breadth
Niche breadth, following Levin's definition, was quantified using Colwell's adjusted formula [32].
where Bi is the niche breadth of the i-th species, Pih is the ratio of the importance value of the i-th species at the h-th resource level to the sum of the importance values of the species at all resource levels, r is the number of resource levels, and the range of the formula is [0,1].
2.5.4. Niche overlap
The Pianka's formula was employed to calculate niche overlap [32].
where Oij is the niche overlap of populations i and j, pih and pij are the proportion of the importance values of the ith and the j-th species at the h-th resource level in the synthesis of the importance values of the species at all resource levels, r is the number of resource levels, and the range of the formula is [0, 1].
2.6. Redundancy Analysis (RDA) and Detrended Correspondence Analysis (DCA)
DCA is a method for sorting and visualizing ecological data distribution. It is usually applied to species abundance data to explore differences between samples or communities. The advantage of DCA is that it can better deal with the nonlinear trends common in ecological data. By removing trends, DCA provides more realistic and interpretable information about the structure of ecological communities. RDA is a multivariate regression technique commonly used to analyze the influence of environmental variables on species composition. It combines the concepts of multiple linear regression and principal component analysis. RDA visualizes the relationship between environmental variables and biological communities by maximizing a linear combination that explains differences in species composition. [34].
RDA and DCA were used in the present study to systematically analyze the species composition, community types and distribution characteristics of vegetation in the two different types of the WLFZs in the Xiangxi River of the TGR, as well as their relationships with environmental factors. We conducted DCA and RDA using the CANOCO package v.5.0 (Ithaca, NY, USA). A Monte Carlo test using 999 permutations (p<0.05) was performed to test the significance of the analysis.
Results
3.1. Plant community characteristics of the RWLFZ and NRZ
A total of 78 vascular plant species, belonging to 62 genus of 24 families, were identified in the Xiangxi River Basin of the TGR (Specific species are shown in Table A1 ( Appendix A )). Of these, 49 vascular plant species of 44 genus and 23 families with 26 endemic species in 22 endemic genus and 8 endemic families were found in the RWLFZs. In the case of the families, few-species and single-species families were dominant, accounting for 61.22% of the total family, and the most abundant species were members of Compositae, followed by Gramineae, which accounted for 24.49% and 14.29% of the total species, respectively. These families contained 33 annual herbs, 13 perennial herbs and 3 trees, accounting for 67.35%, 26.53% and 6.12% of the total species, respectively.
A total of 52 vascular plant species were found in the NRZs, including 13 families and 34 genus (20 endemic species in 18 endemic genus and 3 endemic families), of which 25 annual herbs accounted for 65.79% of all plant species. Both Compositae and Gramineae were the dominant families, both accounting for 23.68% of the plant communities.
3.2. Plant community diversity of the RWLFZs and NRZs
There was no significant difference on Shannon diversity index between NRZs and RWLFZs of the Xiangxi River of the TGR (Figure 3A), but there was significant difference between NRZs and RWLFZs on Pielou evenness index and Simpson dominance index (Figure 3). The result revealed that the Pielou evenness index of NRZs was significantly higher than that of the RWLFZs (Figure 3B), but the Simpson dominance index of RWLFZs was significantly higher than that of NRZs (Figure 3C).
3.3. Functional traits of dominant plant species of the RWLFZs and NRZs
Counter-seasonally hydrological alterations had significant effects on the above-ground biomass (AGB), net photosynthetic rate (Pn), WUE, specific leaf area (SLA) and vegetation coverage (VC) of dominant species in the RWLFZs. Compared with the NRZs, the SLA and VC of the RWLFZs increased by 74.0% and 27% (p<0.05), respectively (Figure 4C,4E), while the WUE significantly decreased by 59.6% (p<0.05) (Figure 4B) and the Pn and AGB between them had no significant difference (Figure 4A,4D). The results showed that counter-seasonally hydrological alterations had the significant effects on functional traits of dominant species in the RWFLZs and dominant species in the RWFLZs grew faster than those in the NRZs with higher SLA and VC, and still maintained high Pn.
3.4. Niche overlap and niche breadth
3.4.1. The reservoir water level fluctuation zones (RWLFZs)
In total, 49 species of herbaceous plants were observed in the RWLFZs. The niche breadth of C. dactylon, B. pilosa, Xanthium sibiricum, Echinochloa crusgalli and Polygonum hydropiper was 0.789, 0.636, 0.505, 0.470 and 0.309, respectively, greater than those of other species. Niche breadth in the RWLFZs varied from 0.0111 to 0.7891. C. dactylon, B. pilosa and X. sibiricum had niche breadths greater than 0.5. Species with niche breadths less than 0.2, accounting for 79.59% of all the species found in the RWLFZs, were mostly distributed in the high elevation areas. And 14 species pairs with niche overlap greater than 0.5 accounted for 17.95% of all species pairs; 22 pairs with niche overlap less than 0.2, accounting for 28.21% of all the species pairs; and 3 pairs with zero niche overlap value, accounting for 3.85% of all the species pairs (Table 2). Among them, the niche overlap value for Eclipta prostrata and Rumex acetosa was the largest (0.712), followed by X. sibiricum (0.662), C. dactylon (0.662), B. pilosa (0.622), Abutilon theophrasti (0.604); P. hydropiper (0.649), E. crusgalli (0.649), Eleusine indica (0.644), E. prostrata and Arthraxon hispidus (0.688). Species pairs with large niche overlap values were mostly those species with large niche breadth and high importance values, but there were also a few species with large niche overlap, low importance values and low niche breadth, such as R. acetosa and A. hispidus (Figure 5A).
3.4.2. The natural riparian zones (NRZs).
There were 38 herbaceous plants in the NRZs, and the niche overlap was calculated for the species pairs with importance values greater than 0.06 (Figure 5B). There were 5 pairs with niche overlap greater than 0.5, accounting for 7.58% of all species pairs; 46 pairs with an overlap of less than 0.2, accounting for 69.7% of all species pairs; and 29 pairs with 0 niche overlap values, accounting for 43.9% of all species pairs. Among them, the highest values were for Conyza canadensis-Artemisia annua (0.922), followed by R. indica-D. orientalis (0.869), B. pilosa-A. annua (0.804), B. pilosa-C. canadensis (0.699), and B. pilosa-R. indica (0.521). Species with large niche overlap values were those species with large niche breadth and high importance values, but C. dactylon, with the highest importance value, and P. hydropiper were not recorded. Among the five pairs of species with niche overlap values greater than 0.5, three were species pairs containing B. pilosa. The number of species with a niche overlap value of 0 in the NRZs was highest in the two zones, accounting for 43.9% of all species pairs in the NRZs. The results showed that the degree of niche differentiation was high in the area.The niche breadth value of the NRZs varied from 0.0833 to 0.648 and was greater than 0.5 for R. indica and B. Pilosa. There were 21 species with a niche breadth of less than 0.2, accounting for 73.68% of all the species found in the NRZs.
3.5. DCA ordination of sampling sites and Species
The DCA ordination results of 105 sampling sites in the Xiangxi River Basin were shown in Figure 6a. On the DCA ordination map, the distribution patterns for each community type were obvious: the vegetation community types in the source area of the NRZ were on the right side of Figure 6a. The distribution patterns of species in the DCA ordination map was coincidently similar to those of community types (Figure 6b). From the horizontal axis of the species DCA ordination map (Figure 5b), at the left end of the horizontal axis, the species were X. sibiricum , C. dactylon , B. pilosa , and A. theophrasti growing in the RWLFZs, while the right end was occupied by typical species such as Debregeasia orientalis, Ricinus communis, and Rorippa indica growing in the NRZs, The variety of species across the distribution reflects the changes in the water level of rhythm characteristics from the NRZs to the RWLFZs of the Xiangxi River Basin. Species at the left end of the horizontal axis, there were mainly flooding-resistant species such as C. dactylon, X. sibiricum, B. pilosa , and A. theophrasti, while at the right end of the horizontal axis mainly included drought-resistant species such as D. orientalis, R. communis, and R. indica. Differences in species composition in different regions indicated the effect of water level rhythms on vegetation.
3.6. RDA of species and environmental variables
Response data were compositional and have a gradient 2.8 SD units long, so linear method was recommended. Therefore, the redundant analysis (RDA) method was used to rank the communities, and the relationship between the distribution pattern of the communities and environmental factors was studied. The ordination analysis revealed the environmental gradient in the area. The results of the RDA showed that elevation was the best explanatory variable (Figure 7). The conditional effects indicated the following:
- (1)
- the contributions of elevation (F=3.3, p <0.01), TK (F=3.2, p <0.01) and flooding duration (F=3.1, p <0.01) were significant during the forward selection; (2) the contributions of the remaining variables were not significant. The RDA results also showed that there was a strong correlation between vegetation and environmental factors, with species-environment correlations of 0.9858 on the first axis and 0.9236 on the second axis. The eigenvalues for the first four RDA axes were 0.2388, 0.128, 0.1133 and 0.0771. The cumulative percentage variance of species-environment relations of the first axis and second axis were 30.42% and 46.73%, respectively. The sum of all the RDA canonical eigenvalues was 0.785. The angles between the vector segments of elevation, TK and flooding duration and the first order axis were small, indicating that these three environmental indicators mainly affected the distribution of species on the first order axis. The arrow of elevation and TK were located in the first quadrant, indicating that elevation and TK was positively related to the first sorting axis. E. villosa, S. viridis and X. sibiricum were distributed in the higher elevation with high TK content. Flooding duration was in the second quadrant and had a negative correlation with the first sort axis. E. indica, P. oleracea and L. procumbens were distributed in the lower elevation. So, the main factors affecting the distribution of the plant community in TGR were elevation, TK and flooding duration.
4. Discussion
4.1. Plant community characteristics and their ecological adaptation strategies in different types of the WLFZs
With the filling of the TGR, original vegetation in the WLFZ between the elevations of 145 and 175 m disappeared and the species richness and diversity significantly decreased due to the great hydrological alterations by the TGD construction [6,35]. The analysis of vegetation responses to river regulation through this dam project is highly important to assess the effects of counter-seasonally hydrological fluctuations on newly-formed riparian ecosystem. In the present study, to better understand the relationships between naturally recovered plant community characteristics and their responses to environmental factors for reconstructing floristically diverse riparian forests, we detected two different types of the WLFZs in the Xiangxi River of the TGR after long-term tracking and monitoring [12,36]. The findings revealed a total of 78 species of vascular plants, belonging to 62 genera and 24 families, within the two distinct types of the WLFZs along the Xiangxi River. Compositae plant accounted for the highest proportion (20.25%), followed by Gramineae (15.38%) and Polygonaceae (7.70%) of all the species detected. There are 7 common families, 15 common genus and 16 common species in the two types of the WLFZs. There were 15 species belonging to one genus of one family, accounting for 19.23% of all the total species. Most of the dominant species were annual herbs in the two different types of the WLFZs after 20 cycles of water level fluctuations and the proportion of the annual dominant species was from 65.79% in the NRZs up to 67.34% in the RWLFZs.
However, differences in plant community composition among the different types of the WLFZs were found. Only 38 of vascular plant species were identified in the NRZs affected by summer floods, but 49 species found in the RWLFZs with the opposite water level rhythm. The taxonomic composition in the RWLFZs was predominantly characterized by the family Compositae, which was the most frequently observed taxon. In contrast, the NRZs exhibited a more balanced distribution, with both Compositae and Gramineae families being equally represented and accounting for the same proportion of the total species count. This distinction in the taxonomic dominance between the two zones highlights the varying ecological preferences and adaptability of these plant families to the specific environmental conditions within the WLFZs. Only 15 species appeared at the same time in the RWLFZs and the NRZs, further highlighting the impact of the anti-seasonally hydrologoical rhythm of the TGR on the vegetation of Xiangxi River. There were 26 endemic species in 22 endemic genus and 8 endemic families in the RWLFZs and 20 endemic species in 18 endemic genus and 3 endemic families in the NRZs. Thus it can be seen that the hydrologoical rhythms of the TGR had the significant effects in plant community composition among different water level fluctuation zones. The dominant species evolved from Cruciferae, Urticaceae, Compositae and Gramineae to Compositae and Gramineae from the NRZs to RWLFZs. The species with the highest importance value in the RWLFZs was C. dactylon (0.4445), followed by C. rotundus (0.1266) and E. crusgalli (0.0965). Perennial plant C. dactylon exhibited an absolute advantage in this type of the RWLFZs. The species with the highest importance value in the NRZs was C. dactylon (0.3884), followed by P. hydropiper (0.1694) and R. indica (0.1585), with the dominant species being members of Gramineae, Polygonaceae and Compositae. In the two different types of the WLFZs, C. dactylon was obviously dominant, showing its stronger ecological adaptability to different environments than other plant species by rapid growth and lateral expansion with seed reproduction and clonal propagation [37,38]. Gramineae species in the RWLFZs demonstrated a robust adaptation to the irregular, anti-seasonal water level fluctuations.. In 2010, Wang et al .[36] found that 64 vascular plant species belonging to 59 genus of 29 families were identified in the same field vegetation survey of the RWLFZs of the Xiangxi River. After seven years following the impoundment of the TGR, Gramineae and Compositae families constituted 17.19% and 14.06% of the total identified plant species, respectively. Fu et al. [12] reported 50 vascular plant species of 49 genus of 23 families occurred in the RWLFZs of the Xiangxi River through the same field study and fixed observation, with Compositae and Gramineae accounting for 16.68% and 13.73%, respectively after 11 years of anti-seasonal flooding. In the present study, a total of 49 vascular plant species were identified, belonging to 44 genus of 23 families in the same studied area of the Xiangxi River with Compositae and Gramineae counting for 20.25% and 15.38%, respectively after 20 years of the impoundment of the TGR. Thus, after experiencing 20 years of off-seasonal water level fluctuations, it was found that the number of plant species considerable decreased in the RWLFZs of the TGR. Compositae plant showed an increasing trend with the number of flood-dry-flood cycles and the number of Compositae has become higher than Gramineae. The reason may be that Compositae has unique nitrogen-carbon balance system,resulting in enhanced N uptake and fatty acid biosynthesis, which may explain their rich biodiversity and excellent adaptive characteristics [65]. Under this type of water level rhythm with high water level in winter and low water level in summer, Compositae and Gramineae plants showed obvious advantages in species number and population size. These results showed if the water-level fluctuation rhythm changes, the composition of annual herb communities will alter because of their rapid responses to the water level fluctuations. A major underlying mechanism which allows annual plants to persist even after years of the water level fluctuations with no reproduction is the maintenance of a persistent seed bank [39]. Therefore, as environmental conditions vary, plant community composition would shift in response to hydrologic changes.
Additionally, in the process of plant community succession, the effects of stress and disturbance on the vegetation assembly result in a particular adaptation strategy of plant species in a particular stage of succession. The competitive (C)-stress tolerant (S)-ruderal (R) (CSR) model of Grime [40] is an attempt to describe the three primary plant adaptation strategies that are linked to the different stages of succession and includes the secondary strategies (CR, SR, SC, and CSR) that correspond to the various trade-offs in the adaptations to competition, stress and disturbance [40~45]. In the present study, the Pn and WUE of dominant plant species from the RWLFZs and NRZs had no significance between them, but the SLA of the former was significantly higher than that of the latter. These indicated the annual species in the RWLFZs adopt the R adaptation strategy with fast-growing, effectively dispersing, short lifespan and quickly acquiring and investing resources by altering leaf morphology such as expanding leaf area into the production of offspring in the primary stage of plant community succession. In habitats with long-term poor conditions, frequent and severe disturbances in the RWLFZs, the resources captured could be quickly reintroduced or redistributed for rapid settlement in the area, annual plants would allocate much more to reproduction in order to be able to maximize the fitness gaining on reproductive output as soon as the environment allows them [46], and plant individuals are relatively small, short life history and high fertility, with annuals germinating in spring and fructifying in autumn or annuals germinating in late winter and fructifying in summer, while in the high elevation areas of the RWLFZs, annual herbs, such as X. sibiricum, grows tall, allocates much more to vertical vegetative growth at the expense of lower reproductive output in order to be able to maximize this species fitness. A progressive replacement of the R-strategists by the SC or the CSR strategists would occur in the long-term succession stages of naturally recovered plant communities, and for perennial plants such as C. dactylon, it adopted the rapid capture and monopolization of resources with the spatially dynamic foraging of roots and shoots according to the C adaptation strategy of the competitive CSR model of Grime. Thus, plant community species at different hydrological regimes would adopt different adaptation strategies in different types of the water level fluctuations.
4.2. Dominant plant niches and biodiversity coexistence and resource utilization mechanism in the different water level fluctuation zones
It is universally acknowledged that the number of related species capable of coexisting in a particular community is contingent upon the niche breadths of individual species as well as the extent to which their niches overlap [47]. The niche is a valid indicator for better understanding of the relationship between species and habitats. It not only reflects the ecological adaptability and distribution range of species but also provides a description of the scarce resources for which species compete [16].Niche breadth reflects the adaptation of species to the environment and the degree of resource utilization. In general, the greater the niche breadth of the species, the stronger is its ability to use resources and adapt to the environment [48]. In the present study, we found the dominant plant species with a niche breadth value of more than 0.5 in the RWLFZs to be C. dactylon, B. pilosa and X. sibiricum. C. dactylon had both the highest importance value and the highest niche breadth value. B. pilosa and X. sibiricum also had relatively high important value and niche breadth value. Species with a niche breadth of less than 0.2 accounted for 79.59% of all the species found in the RWLFZs. However, Wang et al.[49] found that the initial flooding changed the vegetation composition of the original RWLFZs, and many specialized species started to appear and gradually occupied a dominant position (the number of species with a niche breadth less than 0.2 accounting for 90% of all the species pairs) after 5 years of the impoundment in 2008. After 20 years, the proportion of the species with a niche breadth of less than 0.2 (79.59%) in the RWLFZs decreased by approximately 10%, and the dominant position of the specialized species appeared. Although there were only 3 species (C. dactylon, B. pilosa and X. sibiricum) in the region with a niche breadth of more than 0.5, which reflects the stronger ecological adaptability and the ability to use resources, thus quickly expand their own niche space, and the wider distribution range of species in the RWLFZs of the TGR. The NRZ is the zone that is closest to the environment before the construction of the TGR, and the study of the plant communities in this area can highlight changes in vegetation in the Xiangxi River during off-seasonal flooding. The broader niche occupancy, characterized by a wider utilization of environmental resources and spatial domains, typically confers a competitive advantage to certain species. This competitive edge can lead to the exclusion of species with narrower niche widths, thereby facilitating the overall growth and development of the dominant species' guilds within the ecosystem.[66]. Therefore, different species have different ecological adaptation strategies under periodic or aperiodic flooding stress in the different types of WLFZs. In this study, the anti-seasonal flooding rhythm expanded the niche breadth of C. dactylon, B. pilosa, X. sibiricum, E. crusgalli, P. hydropiper and A. theophrasti, showing the ability of these plant species to utilize resources and exhibit high adaptability to dry-wet alternating environments in the RWLFZs. The plant community in the RWLFZs was dominated by the species with the highest or/and second highest niche breadths, revealing that these species had broader environmental tolerances and were able to use a wider range of resources. C. dactylon with the highest important values and niche breadth formed a single-species community in the mid-elevation region of the RWLFZs, and a single species community poses a great threat to the stability of the communities.
Niche overlap is the similarity between two species in relation to ecological factors: when two species use the same resource or share a certain resource, niche overlap occurs [50]. Moreover, niche overlap can to a certain extent reflect the utilization of common resources among populations and also reflect the degree of interlaced distribution between species [48]. Under the effect of long-term and anti-seasonal water level fluctuations of the TGR, the dry-wet alternating process leads to the surviving resource scarcity of species.Li et al.[67]found that the soil nutrient content decreased significantly after dry-wet alternation. Plants need to expand their resource utilization scope to increase their survival opportunities in unstable habitats to promote niche differentiation and increase niche breadth and niche overlap value. Although the niche breadth of species in the RWLFZs was generally small, the overall niche overlap was relatively high, that is, species competition in the habitat was intense, which was not conducive to the coexistence of species, and the community structure was unstable. Based on the comprehensive analysis of the overlap of plant niches in the two different WLFZs, it can be inferred that after experiencing anti-seasonal flooding, the overlap of niches between all species in the RWLFZs was relatively high, which showed that the competition in the community was much more intraspecific than interspecific and then would facilitate the expansion of species niches. This is consistent with the conclusions drawn by Wang et al. [68] who found that "as the number of reservoir water level fluctuation cycles increases, the niche width and niche overlap of dominant plant populations in the drawdown zone will increase." The reason for this may be the impact of habitat stability. The habitat of the drawdown zone is constantly changing, and in order to gain an advantage under habitat pressure and competition, plant populations tend to expand their range of resource utilization, thus increasing the niche width. Consequently, the likelihood of plant populations utilizing similar resources increases, and the proportion of niche similarity and the value of niche overlap are correspondingly enhanced [69].
However, the competition in the community at the NRZs was much more interspecific than intraspecific. The intensive interspecies competition not only excluded some species with less adaptive faculty but also segregated and optimized niches of the remaining species, leading to a decline in total niche overlap of all paired species in the community. The extent of interspecific competition in the community was also weakened and most species could utilize the environmental resources more coordinately and adequately, which showed that the community structure in the NRZs was extremely stable.
The species of coexistence not only compete for light, heat, water and nutrient resources, but also has obvious facilitation effect, especially in conjunction with high environmental stress [51]. The biggest problem under flooding stress is the reduction of oxygen supply, so plants have to switch from aerobic respiration to anaerobic respiration and aggravate the consumption of nutrients. Some flood-tolerant wetland plants can alleviate soil anoxia by releasing oxygen into the soil in flooded environments, simultaneously, neighboring flood-sensitive species should grow well under high water levels because flood-tolerant plants ameliorate soil anoxia-stress [52]. In fact, the improvement of anoxic environment depends mainly on the roots of the plant. According to the survey, the C. dactylon is a dominant plant species in different altitude gradients of the RWFLZs and NRZs, this is mainly due to the larger root porosity and the effective scavenging mechanism of reactive oxygen from C. dactylon under inundation conditions. Different species adopt different strategies to cope with the flooding-stress. Some species with high flood tolerance can promote the growth of stems and leaves to achieve higher biomass through the elongation of stems and the leaf area expansion. It facilitates the formation of a better living environment to achieve coexistence [53].
4.3. Distribution patterns of species diversity in the RWLFZ
Environmental gradients provide a useful framework for evaluating the relative importance of differential selection for adaptive traits maximizing species fitness [54,55]. In the present study, an intriguing consequence of our study was that there was no significant difference between NRZs and RWLFZs on Shannon diversity index (Figure 3A), but there is significant difference between NRZs and RWLFZs on Pielou evenness index and Simpson dominance index (Figure 3). The Pielou evenness index of NRZs was significantly higher than that of RWLFZs (Figure 3B), while the Simpson dominance index of RWLFZs was significantly higher than that of NRZs (Figure 3C), which showed that species patterns of plant communities in the RWLFZs were more uneven than in the NRZs and there are more specialized species in the RWLFZs than in the NRZs. The diversity evidences further confirmed the plant community structure in the RWLFZs was still unstable.
The distribution patterns of plant community species diversity along an elevation gradient and in different habitats may be influenced by environmental factors (e.g. latitude and longitude, elevation, flooding time, flooding depth, slope, aspect, human activity) [56]. Chen et al. [11] reported that the steep slope area above 25° in the Xiangxi River of TGR accounted for 41.19% of the RWLFZs, which belongs to the semi-submerged and semi-exposed type of the RWLFZs, especially often under high temperature and drought stress in summer, and the soil organic matter is not readily retained (Figure 2), resulting in a slight decrease in species diversity in the RWLFZs. And all soil nutrients measured also showed in an inverted hump-shaped pattern, except TK in the mid-elevation area. Species with short life history like E. indica, P. oleracea and L. procumbens, were associated with flooding duration in the RDA, whereas tall and loosely branched species, like E. villosa, S. viridis and X. sibiricum, indeed showed a negative relationship to flooding duration (Figure 7) in the RWLFZs. The left to the right ordination diagram of the horizontal axis of RDA showed from the low elevation to the high elevation, plant community architecture altered from small, compact plants in the low-elevation areas to tall, loosely branched plants in the high-elevation areas, where biomass, height and density of neighbors were relatively high. The distribution trend was basically consistent with its dominant species community. The results of the Monte Carlo random displacement test showed that the simple term effect of interpretation of species distribution on elevation, TK and flooding duration reached a significant level. The concentration of total K increased from 2008 to 2016, while the concentrations of OM and available K declined over the same period in the TGR [57]. K can regulate water and energy metabolism and improve plant resistance [58,59]. The close correlation between TK and elevation explained the trend of community diversity. Therefore, the main environmental constraint is most likely elevation, TK and flooding duration. The environment factors drove the succession of vegetation. Vegetation also plays an important role in nutrient cycling directly via plant uptake, decomposition, and deposition of alluvial materials, and indirectly through stimulation of microbial processes in the rhizosphere [57]. The synergy between environmental factors and vegetation played an important role in the natural vegetation restoration in the TGR.
In the study of long-term responses of the WLFZ vegetation to water-level regulation, Nilsson et al.[10,60,61,62] assessed the long-term effect of water-level regulation on riparian plant communities for storage reservoirs and run-of-river impoundments in central and northern Sweden after regulation. The results showed that transformed species richness in pre-upland sites along storage reservoirs gradually increased during the first 34 years but decreased thereafter between 1 and 70 years after dam closure. Along run-of-river impoundments, pre-upland sites had maximum species richness at about 13 years, while pre-riparian sites had maximum species richness at about 18 years after regulation. In the present study, after the first impoundment in June 2003, the TGR has experienced 20 times of the water level fluctuation events, species composition was single, community structure was also simple, the niche differentiation between different dominant plants was lower, and the inter-specific competition was more intense.
4.4. Restoration implications
The new insight on the vegetation distribution patterns of NRZs and RWLFZs through plant niches analyses showed that the inter-specific competition of vegetation distribution in the RWLFZs was more intense due to repeated anti-seasonal flooding stress compared with the NRZs. Plant community architecture altered from small, compact plants in the low-elevation areas to tall, loosely branched plants in the high-elevation areas, where biomass, height and density of neighbors were also relatively high, and C. dactylon, B. pilosa, X. sibiricum, P. hydropiper, S. viridis, E. crusgalli had larger niche breadths, their resource utilization and environmental adaptability were strong, and had both ecological and economic values. Accordingly, these species should be prioritized for optimal selection and scientific arrangement in the ecological restoration and reconstruction of the vegetation in the RWLFZs of the TGR. Moreover, annual herbaceous plants are better suited to the fragmented conditions of the RWLFZs compared to perennial herbaceous species. It is therefore recommended that the general policy for restoring and establishing artificial plant communities in the reservoir area should primarily consist of herbaceous species, complemented by flooding-tolerant shrubs or small trees such as Distylium chinense [63].
In order to establish a complete reference system for vegetation restoration, restoration monitoring plots in the different succession stages should be established in the different water level fluctuation zones of the TGR, and their environmental conditions, community structures and inter-specific relationships should also be analyzed. Therefore, our findings are useful for anticipating the highly adaptable and widely distributed species with large niche breadths and high important values selected as the candidate species in ecological restoration and revegetation in the newly formed hydro-fluctuation zone and the other similar degenerative riparian area and stream ecosystems [70].
5. Conclusion
A total of 76 species of vascular plants were found in the WLFZs of the Xiangxi River Basin. There were 49 and 38 species in the RWLFZs and NRZs, respectively. There were 26 endemic species in 22 endemic genus and 8 endemic families in the RWLFZs and 20 endemic species in 18 endemic genus and 3 endemic families in the NRZs. The dominant species belonged to Cruciferae, Urticaceae, Compositae and Gramineae to Compositae and Gramineae in the NRZs to the RWLFZs.The analysis of niche overlap revealed the instability of the plant community in the RWLFZs, which was attributed to intense intraspecific competition and incomplete niche differentiation. Specifically, species with larger niche widths in the RWLFZs exhibited a high degree of niche overlap, indicating the intensity of resource competition within the plant community in this area. In contrast, the plant communities in the NRZs show lower levels of niche overlap, reflecting greater niche differentiation and a more stable community structure. The results of the RDA indicated that the main factors influencing the vegetation characteristics were the elevation (flooding depth), TK and flooding duration. Our results had some implications for the vegetation restoration efforts of the RWLFZs.The highly adaptable and widely distributed species with large niche breadths and high important values could be selected as the candidate species in ecological restoration and revegetation in the newly formed hydro-fluctuation zones and the other similar degenerative riparian area and stream ecosystems.
Author Contributions
Conceptualization, X.L. and Z.Y.; methodology,D.D.; software, S.X. and W.Y.; validation, D.H.,Q.M. and G.C.; formal analysis, W.Y.; investigation, J.Y.; resources, G.H.; data curation, M.H.; writing—original draft preparation, D.H. and S.X.; writing—review and editing, W.Y. and C.Y.; visualization, X.L. and Z.Y.; supervision, X.L. and C.Y.; project administration, X.L.; funding acquisition, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 51779127), partially funded by National Key R&D Program of China (2023YFF1305204), Research Project on Seed Preservation Technology and Facilities of Rare Plants in the Three Gorges Reservoir Area—Investigation and Collection of Flooding-Tolerant Germplasm Resources (SDHZ2021346) and the 111 Project of China (No. D20015).
Data Availability Statement
The datasets used in this study can be obtained by contacting the corresponding author upon request.
Acknowledgments
Great appreciation is extended to Lei Sun, Lei Yue, Ling Xiang, Jie Huang and Cheng-Rui Yu for their assistance during field sampling and laboratory analysis. We would like to express our gratitude to the anonymous reviewers for their insightful feedback and constructive suggestions, which significantly enhance the quality of this manuscript.
Conflicts of Interest
The authors declared that they have no conflictsof interest to this work.
Appendix A

Table A1.
Changes of composition and niche breadth of dominant herbaceous plants.
| Species | Life form | Family | Genera | ||||
| WLFZ | NRZ | ||||||
| Lv | Bi | Lv | Bi | ||||
| Cynodon dactylon | Perennial herb | Gramineae | Cynodon | 0.789 | 0.444 | 0.478 | 0.289 |
| Bidens pilosa | Annual | Compositae | Bidens | 0.636 | 0.059 | 0.461 | 0.076 |
| herbaceous | |||||||
| Xanthium sibiricum | Annual | Compositae | Xanthium | 0.505 | 0.068 | 0.228 | 0.101 |
| herbaceous | |||||||
| Echinochloa crusgalli | Annual | Gramineae | Echinochloa | 0.47 | 0.097 | 0.371 | 0.193 |
| herbaceous | |||||||
| Polygonum hydropiper | Annual | Polygonaceae | Polygonum | 0.309 | 0.065 | 0.197 | 0.088 |
| herbaceous | |||||||
| Abutilon theophrasti | Annual | Malvaceae | Abutilon | 0.305 | 0.061 | 0.48 | 0.047 |
| herbaceous | |||||||
| Acalypha australis | Annual | Euphorbiaceae | Acalypha | 0.293 | 0.028 | 0.056 | 0.008 |
| herbaceous | |||||||
| Eclipta prostrata | Annual | Eclipta | 0.28 | 0.056 | 0.465 | 0.065 | |
| herbaceous | Compositae | ||||||
| Cyperus rotundus | Perennial herb | Cyperaceae | Cyperus | 0.249 | 0.127 | 0.1 | 0.073 |
| Setaria viridis | Annual | Gramineae | Setaria | 0.24 | 0.078 | 0.376 | 0.112 |
| herbaceous | |||||||
| Digitaria sanguinalis | Annual | Gramineae | Digitaria | 0.131 | 0.041 | 0.192 | 0.021 |
| herbaceous | |||||||
| Eleusine indica | Annual | Gramineae | Eleusine | 0.129 | 0.085 | 0.289 | 0.057 |
| herbaceous | |||||||
| Polygonum lapathifolium | Annual | Polygonaceae | Polygonum | 0.125 | 0.023 | - | - |
| herbaceous | |||||||
| Arthraxon hispidus | Annual | Gramineae | Arthraxon | 0.108 | 0.023 | - | - |
| herbaceous | |||||||
| Phyllanthus urinaria | Annual | Euphorbiaceae | Phyllanthus | 0.089 | 0.02 | - | - |
| herbaceous | |||||||
| Amaranthus retroflexus | Annual | Amaranthaceae | Amaranthus | 0.085 | 0.042 | 0.13 | 0.038 |
| herbaceous | |||||||
| Portulaca oleracea | Annual | Portulacacea | Portulaca | 0.085 | 0.037 | 0.111 | 0.01 |
| herbaceous | |||||||
| Eriochloa villosa | Annual | Gramineae | Eriochloa | 0.08 | 0 | - | - |
| herbaceous | |||||||
| Conyza canadensis | Annual | Compositae | Conyza | 0.077 | 0.014 | 0.212 | 0.053 |
| herbaceous | |||||||
| Solanum nigrum | Annual | Solanaceae | Solanum | 0.076 | 0.017 | 0.143 | 0.072 |
| herbaceous | |||||||
| Alternanthera philoxeroides | Perennial herb | Amaranthaceae | Alternanthera | 0.071 | 0.02 | - | - |
| Pharbitis purpurea | Annual | Convolvulaceae | Pharbitis | 0.049 | 0.121 | - | - |
| herbaceous | |||||||
| Artemisia argyi | Perennial herb | Compositae | Artemisia | 0.044 | 0.013 | 0.124 | 0.235 |
| Humulus scandens | Annual twining herb | Moraceae | Humulus | 0.042 | 0.045 | 0.148 | 0.021 |
| Lindernia procumbens | Annual | Scrophulariaceae | Lindernia | 0.041 | 0.026 | 0.102 | 0.02 |
| herbaceous | |||||||
| Artemisia capillaris | Perennial herb | Compositae | Artemisia | 0.041 | 0.017 | - | - |
| Mazus japonicus | Annual | Scrophulariaceae | Mazus | 0.035 | 0.031 | - | - |
| herbaceous | |||||||
| Gnaphalium affine | Annual | Compositae | Gnaphalium | 0.033 | 0.005 | - | - |
| herbaceous | |||||||
| Crassocephalum crepidioides | Annual | Compositae | Crassocephalum | 0.033 | 0.01 | 0.083 | 0.034 |
| herbaceous | |||||||
| Kummerowia striata | Annual | Leguminosae | Kummerowia | 0.033 | 1.887 | 0.138 | 0.015 |
| herbaceous | |||||||
| Daucus carota | Perennial herb | Umbelliferae | Daucus | 0.025 | 0.012 | - | - |
| Phytolacca acinosa | Perennial herb | Phytolaccaceae | Phytolacca | 0.022 | 0.046 | - | - |
| Chenopodium album | Annual | Chenopodiaceae | Chenopodium | 0.022 | 0.053 | - | - |
| herbaceous | |||||||
| Siegesbeckia orientalis | Annual | Compositae | Siegesbeckia | 0.021 | 0.038 | - | - |
| herbaceous | |||||||
| Polygonum plebeium | Annual | Polygonaceae | Polygonum | 0.019 | 0.005 | 0.037 | 0.006 |
| herbaceous | |||||||
| Stellaria media | Biennial herbs | Caryophyllaceae | Stellaria | 0.011 | 0.002 | 0.083 | 0.041 |
| Gynura formosana | Perennial herb | Compositae | Gynura | 0.011 | 0.004 | - | - |
| Clinopodium chinense | Perennial herb | Labiatae | Clinopodium | 0.011 | 0.002 | - | - |
| Sapium sebiferum | Tree | Euphorbiaceae | Sapium | 0.011 | 0.004 | - | - |
| Convolvulus arvensis | Perennial herb | Convolvulaceae | Convolvulus | 0.011 | 0.003 | - | - |
| Paederia scandens | Perennial twining herb | Rubiaceae | Paederia | 0.011 | 0.005 | - | - |
| Lindernia procumbens | Annual | Scrophulariaceae | Lindernia | 0.011 | 0.011 | - | - |
| herbaceous | |||||||
| Cucumis sativus | Annual Vines herb | Cucurbitaceae | Cucumis | 0.011 | 0.003 | - | - |
| Melilotus officinalis | Biennial herbs | Leguminosae | Melilotus | 0.011 | 0.039 | - | - |
| Dendranthema indicum | Perennial herb | Compositae | Dendranthema | 0.011 | 0.009 | - | - |
| Broussonetia papyrifera | Tree | Moraceae | Broussonetia | 0.011 | 0.024 | - | - |
| Corydalis pallida | Annual | Papaveraceae | Corydalis | 0.011 | 0.019 | - | - |
| herbaceous | |||||||
| Ixeridium chinense | Perennial herb | Compositae | Ixeridium | 0.011 | 0.031 | - | - |
| Rhus chinensis | Deciduous small trees or shrubs | Anacardiaceae | Rhus | 0.011 | 0.071 | - | - |
| Pennisetum alopecuroides | Perennial herb | Gramineae | Pennisetum | - | - | 0.184 | 0.035 |
| Chenopodium ambrosioides | Annual or perennial herbs | Chenopodiaceae | Chenopodium | - | - | 0.101 | 0.008 |
| Salix variegata | shrub | Salicaceae | Salix | - | - | 0.093 | 0.014 |
| Ludwigia prostrata | Annual | Onagraceae | Ludwigia | - | - | 0.093 | 0.017 |
| herbaceous | |||||||
| Cyperus difformis | Annual | Cyperaceae | Cyperus | - | - | 0.12 | 0.016 |
| herbaceous | |||||||
| Artemisia annua | Annual | Compositae | Artemisia | - | - | 0.177 | 0.04 |
| herbaceous | |||||||
| Ranunculus sieboldii | Perennial herb | Ranunculaceae | Ranunculus | - | - | 0.056 | 0.005 |
| Oxalis corniculata | Perennial herb | Oxalidaceae | Oxalis | - | - | 0.037 | 0.008 |
| Youngia japonica | Annual | Compositae | Youngia | - | - | 0.098 | 0.047 |
| herbaceous | |||||||
| Rorippa indica | Biennial herbs | Cruciferae | Rorippa | - | - | 0.648 | 0.158 |
| Eragrostis minor | Annual | Poaceae | Eragrostis | - | - | 0.246 | 0.024 |
| herbaceous | |||||||
| Oxalis corniculata | Annual | Oxalidaceae | Oxalis | - | - | 0.167 | 0.025 |
| herbaceous | |||||||
| Debregeasia orientalis | Deciduous small trees or shrubs | Urticaceae | Debregeasia Gaudich. | - | - | 0.496 | 0.098 |
| Veronica didyma | Biennial herbs | Scrophulariaceae | Veronica L. | - | - | 0.083 | 0.012 |
| Cerastium arvense | Perennial herb | Caryophyllaceae | Cerastium | - | - | 0.162 | 0.016 |
| Arabidopsis thaliana | Annual or perennial herbs | Cruciferae | Arabidopsis | - | - | 0.083 | 0.066 |
| Cardamine hirsuta | Annual | Cruciferae | Cardamine | - | - | 0.166 | 0.039 |
| herbaceous | |||||||
| Fimbristylis dichotoma | Annual | Cyperaceae | Fimbristylis Vahl | - | - | 0.083 | 0.06 |
| herbaceous | |||||||
| Buddleja lindleyana | shrub | Loganiaceae | Buddleja | - | - | 0.167 | 0.036 |
| Rumex japonicus | Perennial herb | Polygonaceae | Rumex | - | - | 0.083 | 0.016 |
| Rumex acetosa | Perennial herb | Polygonaceae | Rumex | - | - | 0.083 | 0.01 |
| Polygonum capitatum | Perennial herb | Polygonaceae | Polygonum | - | - | 0.083 | 0.011 |
| Ambrosia artemisiifolia | Annual or perennial herbs | Compositae | Ambrosia | - | - | 0.083 | 0.054 |
| Galinsoga parviflora | Annual | Compositae | Galinsoga | - | - | 0.25 | 0.051 |
| herbaceous | |||||||
| Polypogon fugax | Annual | Gramineae | Polypogon | - | - | 0.083 | 0.04 |
| herbaceous | |||||||
| Cenchrus echinatus | Annual | Gramineae | Cenchrus | - | - | 0.152 | 0.048 |
| herbaceous | |||||||
| Brachypodium sylvaticum | Perennial herb | Gramineae | Brachypodium | - | - | 0.083 | 0.017 |
| Vicia sepium | Perennial herb | Leguminosae | Vicia | - | - | 0.083 | 0.009 |
| Ricinus communis | Annual | Euphorbiaceae | Ricinus | - | - | 0.083 | 0.033 |
Table A2.
Each number in Figure 6b represents the species.
Table A2.
Each number in Figure 6b represents the species.
| Number | Plant species | Number | Plant species |
|---|---|---|---|
| 1 | Echinochloa crusgalli | 36 | Chenopodium album |
| 2 | Polygonum hydropiper | 37 | Humulus scandens |
| 3 | Portulaca oleracea | 38 | Melilotus officinalis |
| 4 | Cynodon dactylon | 39 | Artemisia capillaris |
| 5 | Eleusine indica | 40 | Cucumis sativus |
| 6 | Arthraxon hispidus | 41 | Eriochloa villosa |
| 7 | Cyperus rotundus | 42 | Kummerowia striata |
| 8 | Eclipta prostrata | 43 | Daucus carota |
| 9 | Amaranthus retroflexus | 44 | Dendranthema indicum |
| 10 | Polygonum plebeium | 45 | Corydalis pallida |
| 11 | Rumex acetosa | 46 | Broussonetia papyrifera |
| 12 | Lindernia procumbens | 47 | Pennisetum alopecuroides |
| 13 | Abutilon theophrasti | 48 | Artemisia annua |
| 14 | Bidens pilosa | 49 | Oxalis corniculata |
| 15 | Mazus japonicus | 50 | Youngia japonica |
| 16 | Phyllanthus urinaria | 51 | Rumex japonicus |
| 17 | Phytolacca acinosa | 52 | Polygonum lapathifolium |
| 18 | Xanthium sibiricum | 53 | Rorippa indica |
| 19 | Crassocephalum crepidioides | 54 | Arabidopsis thaliana |
| 20 | Acalypha australis | 55 | Veronica didyma |
| 21 | Gnaphalium affine | 56 | Fimbristylis dichotoma |
| 22 | Solanum nigrum | 57 | Debregeasia orientalis |
| 23 | Stellaria media | 58 | Vicia sepium |
| 24 | Gynura formosana | 59 | Ricinus communis |
| 25 | Setaria viridis | 60 | Buddleja lindleyana |
| 26 | Alternanthera philoxeroides | 61 | Ambrosia artemisiifolia |
| 27 | Clinopodium chinense | 62 | Galinsoga parviflora |
| 28 | Digitaria sanguinalis | 63 | Polypogon fugax |
| 29 | Sapium sebiferum | 64 | Cenchrus echinatus |
| 30 | Convolvulus arvensis | 65 | Polygonum capitatum |
| 31 | Pharbitis purpurea | 66 | Brachypodium sylvaticum |
| 32 | Siegesbeckia orientalis | 67 | Cerastium arvense |
| 33 | Artemisia argyi | 68 | Eragrostis minor |
| 34 | Conyza canadensis | 69 | Cardamine hirsuta |
| 35 | Paederia scandens |
Table A3.
Each number in Figure 7 represents the species.
Table A3.
Each number in Figure 7 represents the species.
| Number | Plant species | Number | Plant species |
|---|---|---|---|
| 1 | Cynodon dactylon | 26 | Artemisia capillaris |
| 2 | Bidens pilosa | 27 | Mazus japonicus |
| 3 | Xanthium sibiricum | 28 | Gnaphalium affine |
| 4 | Echinochloa crusgalli | 29 | Crassocephalum crepidioides |
| 5 | Polygonum hydropiper | 30 | Kummerowia striata |
| 6 | Abutilon theophrasti | 31 | Daucus carota |
| 7 | Acalypha australis | 32 | Phytolacca acinosa |
| 8 | Eclipta prostrata | 33 | Chenopodium album |
| 9 | Cyperus rotundus | 34 | Siegesbeckia orientalis |
| 10 | Setaria viridis | 35 | Polygonum plebeium |
| 11 | Digitaria sanguinalis | 36 | Stellaria media |
| 12 | Eleusine indica | 37 | Gynura formosana |
| 13 | Polygonum lapathifolium | 38 | Clinopodium chinense |
| 14 | Arthraxon hispidus | 39 | Sapium sebiferum |
| 15 | Phyllanthus urinaria | 40 | Convolvulus arvensis |
| 16 | Amaranthus retroflexus | 41 | Paederia scandens |
| 17 | Portulaca oleracea | 42 | Lindernia procumbens |
| 18 | Eriochloa villosa | 43 | Cucumis sativus |
| 19 | Conyza canadensis | 44 | Melilotus officinalis |
| 20 | Solanum nigrum | 45 | Dendranthema indicum |
| 21 | Alternanthera philoxeroides | 46 | Broussonetia papyrifera |
| 22 | Pharbitis purpurea | 47 | Corydalis pallida |
| 23 | Artemisia argyi | 48 | Ixeridium chinense |
| 24 | Humulus scandens | 49 | Rhus chinensis |
| 25 | Cenchrus echinatus |
References
- Benchimol, M.; Peres, C.A. Predicting local extinctions of Amazonian vertebrates in forest islands created by a mega dam. Biol. Conserv. 2015, 187, 61–72. [Google Scholar] [CrossRef]
- Sun, L.; Ding, W.; Zhou, Y.; Wang, J.; Ouyang, X.; Fan, Z.; Yao, Y.; Zhang, C. Tree Radial Growth Responses to Climate and Reservoir Impoundment in Valleys in SouthwesternChina. Forests 2024, 15, 749. [Google Scholar] [CrossRef]
- Barnes, B.; Sidhu, H.; Roxburgh, S. A model integrating patch dynamics, competing species and the intermediate disturbance hypothesis. Ecol. Modell. 2006, 194, 414–420. [Google Scholar] [CrossRef]
- Ali, M.M. Shoreline vegetation of Lake Nubia, Sudan. In Proceedings of the Macrophytes in Aquatic Ecosystems: From Biology to Management:Proceedings of the 11th Internatonal Symposium on Aquatic Weeds, European Weed Research Society; 2006; pp. 101–105. [Google Scholar] [CrossRef]
- Zhang, Z.; Wan, C.; Zheng, Z.; Hu, L.; Feng, K.; Chang, J.; Xie, P. Plant community characteristics and their responses to environmental factors in the water level fluctuation zone of the three gorges reservoir in China. Environ. Sci. Pollut. Res. 2013, 20, 7080–7091. [Google Scholar] [CrossRef]
- Ye, C.; Zhang, K.; Deng, Q.; Zhang, Q. Plant communities in relation toflooding and soilcharacteristics in the water level fluctuation zone of the Three Gorges Reservoir, China. Environ. Sci. Pollut. Res. 2013, 20, 1794–1802. [Google Scholar] [CrossRef]
- Bao, Y.; Gao, P.; He, X. The water-level fluctuation zone of Three Gorges Reservoir—A unique geomorphological unit. Earth-Sci. Rev. 2015, 150, 14–24. [Google Scholar] [CrossRef]
- Naiman, R.J.; Melillo, J.M.; Lock, M.A.; Ford, T.E.; Reice, S.R. Longitudinal patterns of ecosystem processes and community structure in a subarctic river continuum. Ecology 1987, 68, 1139–1156. [Google Scholar] [CrossRef]
- Jansson, R.; Nilsson, C.; Dynesius, M.; Andersson, E. Effects of river regulation on river-margin vegetation: a comparison of eight boreal rivers. Ecol. Appl. 2000, 10, 203–224. [Google Scholar] [CrossRef]
- Nilsson, C.; Jansson, R. Floristic differences between riparian corridors of regulated and free-flowing boreal rivers. Regul. Rivers: Res. Manage. 1995, 11, 55–66. [Google Scholar] [CrossRef]
- Chen, Z.L.; Yuan, X.Z.; Liu, H.; Li, B. Characteristics of plant communities in the hydrofluctuation zones of the TGR area under water level fluctuation. Resour. Environ. Yangtze Basin, 2012; 21, 672–677. [Google Scholar]
- Yuan, S.; Zeng, B.; Su, X.; Xu, J. Effect of water-level fluctuation discrepancy on the composition of different annuals in Three Gorges reservoir drawdown zone. Acta Ecol. Sin. 2014, 34, 6481–6488. [Google Scholar]
- Su, X.; Bejarano, M.D.; Yi, X.; Lin, F.; Ayi, Q.; Zeng, B. Unnatural flooding alters the functional diversity of riparian vegetation of the Three Gorges Reservoir. Freshwater Biol. 2020, 65, 1585–1595. [Google Scholar] [CrossRef]
- Noble, R.E.; Murphy, P.K. Short term effects of prolonged backwater flooding on understory vegetation. Castanea, 1975; 228–238. [Google Scholar]
- Yuan, S.; Zeng, B.; Su, X.; Xu, J. Effect of water-level fluctuation discrepancy on the composition of different annuals in Three Gorges reservoir drawdown zone. Acta Ecol. Sin. 2014; 34, 6481–6488. (in Chinese) [Google Scholar]
- Guo,Y.; Yang, S.; Shen, Y.F.; Xiao, W.F.; Rui, R.M. Composition and niche of the existing herbaceous plants in the water-level-fluctuating zone of the TGR Area, China. Chin. J. Appl. Ecol.2018, 29, 3559-3568. 29. [CrossRef]
- Guo, Q.; Kang, Y.; Hong, M.; Jin, J.; Zhu, N.; Nie, B.; Wang, Z. Responses of terrestrial plants in hydro-fluctuation belt of the Three Gorges Reservoir Area to the first time flooding-drying habitat change. Sci. Silvae Sin. 2013, 49, 1–9. [Google Scholar] [CrossRef]
- Zhang, A.; Fan, D.; Li, Z.; Xiong, G.; Xie, Z. Enhanced photosynthetic capacity by perennials in the riparian zone of the Three Gorges Reservoir Area, China. Ecol. Eng. 2016, 90, 6–11. [Google Scholar] [CrossRef]
- Harrison, M.T.; Kelman, W.M.; Moore, A.D.; Evans, J.R. Grazing winter wheat relieves plant water stress and transiently enhances photosynthesis. Funct. Plant Biol. 2010, 37, 726–736. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Levin, S.A.; Root, R.B. Niche, habitat, and ecotope. Am. Nat. 1973, 107, 321–338. [Google Scholar] [CrossRef]
- Petraitis, P.S. Likelihood measures of niche breadth and overlap. Ecology 1979, 60, 703–710. [Google Scholar] [CrossRef]
- Ganis, P. NICHE—Programs for Niche Breadth, Overlap and Hypervolumes. In Computer ssisted egetation analysis; Springer: 1991; pp. 469–487. [CrossRef]
- Slatyer, R.A.; Hirst, M.; Sexton, J.P. Niche breadth predicts geographical range size: a general ecological pattern. Ecol. Lett. 2013, 16, 1104–1114. [Google Scholar] [CrossRef]
- Malkinson, D.; Tielbörger, K. What does the stress-gradient hypothesis predict? Resolving the discrepancies. Oikos 2010, 119, 1546–1552. [Google Scholar] [CrossRef]
- Yan, H.; Huang, Y.; Wang, G.; Zhang, X.; Shang, M.; Feng, L.; Dong, J.; Shan, K.; Wu, D.; Zhou, B. Water eutrophication evaluation based on rough set and petri nets: A case study in Xiangxi-River, Three Gorges Reservoir. Ecol. Indic. 2016, 69, 463–472. [Google Scholar] [CrossRef]
- Maloof, J.N.; Nozue, K.; Mumbach, M.R.; Palmer, C.M. LeafJ: an ImageJ plugin for semi-automated leaf shape measurement. J. Visualized Exp. 2013, e50028. [Google Scholar] [CrossRef]
- Wu, X.; Fan, W.; Du, H.; Ge, H.; Huang, F.; Xu, X. Estimating crown structure parameters of moso bamboo: Leaf area and leaf angle distribution. Forests 2019, 10, 686. [Google Scholar] [CrossRef]
- Kabała, C.; Musztyfaga, E.; Gałka, B.; Łabuńska, D.; Mańczyńska, P. Conversion of Soil pH 1: 2.5 KCl and 1: 2.5 H2O to 1: 5 H 2 O: Conclusions for Soil Management, Environmental Monitoring, and International Soil Databases. Pol. J. Environ. Stud, 2016; 25. [Google Scholar] [CrossRef]
- Forch, J. A simple method for the isolation and purification of total lipids from animal tissues. J. biol. Chem. 1957, 226, 498–509. [Google Scholar] [CrossRef]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. Mod. Methods Plant Anal. 1960, 5, 3–5. [Google Scholar]
- Ye, C.; Li, S.; Zhang, Y.; Zhang, Q. Assessing soil heavy metal pollution in the water-level-fluctuation zone of the Three Gorges Reservoir, China. J. Hazard. Mater. 2011, 191, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Tan, L. Statistical Ecology. China Forestry Press: Beijing, China, 2009.
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Takane, Y.; Hwang, H. Regularized linear and kernel redundancy analysis. Comput Stat. Data. An 2007, 52, 394–405. [Google Scholar] [CrossRef]
- Zhu, K.W.; Chen, Y.C.; Zhang, S.; Lei, B.; Yang, Z.M.; Huang, L. Vegetation of the water-level fluctuation zone in the Three Gorges Reservoir at the initial impoundment stage. Global Ecol. Conserv. 2020, 21, e00866. [Google Scholar] [CrossRef]
- Wang, F.; Xu, T.; Huang, Y.P. Investigation on plant community and distribution characteristics of Xiangxi River bank. Int. J. Green Technol. 2014, 4. [Google Scholar]
- Xu, Y.; Chena, F. Effects of concrete content in vegetation concrete matrix on seed germination and seeding establishment of cynodon dactylon. Procedia Eng. 2012, 28, 105–109. [Google Scholar] [CrossRef]
- Singh, K.; Pandey, V.C.; Singh, R.P. Cynodon dactylon: an efficient perennial grass to revegetate sodic lands. Ecol. Eng. 2013, 54, 32–38. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Xiong, G. M. ; Fan. D.Y.; Xie, Z.Q.Effects of damming on plant diversity in the inundated and riparian zones of the TGR Area, China. Chin. /: https, 2016; ,35. [Google Scholar]
- Grime, J.P. Plant strategies, vegetation processes, and ecosystem properties. 2001.
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grime, J. Dominant and subordinate components of plant communities: implications for succession, stability and diversity. In Proceedings of the Symposium of the British Ecological Society; 1987. [Google Scholar]
- Liao, J.; Jiang, M.; Li, L. Effects of simulated submergence on survival and recovery growth of three species in water fluctuation zone of the Three Gorges reservoir. Acta Ecol. Sin. 2010, 30, 216–220. [Google Scholar] [CrossRef]
- Chai, Y.; Yue, M.; Wang, M.; Xu, J.; Liu, X.; Zhang, R.; Wan, P. Plant functional traits suggest a change in novel ecological strategies for dominant species in the stages of forest succession. Oecologia 2016, 180, 771–783. [Google Scholar] [CrossRef]
- Yi, X.; Huang, Y.; Ma, M.; Wen, Z.; Chen, J.; Chen, C.; Wu, S. Plant trait-based analysis reveals greater focus needed for mid-channel bar downstream from the Three Gorges Dam of the Yangtze River. Ecol. Indic. 2020, 111, 105950. [Google Scholar] [CrossRef]
- Pisula, W.; Łukasik, A.; Kawa, R.; Pisula, E. Exploratory analysis of the links among life history, reproductive strategy, autism-spectrum quotient, and quality of life. Psychology 2018, 9, 2312. [Google Scholar] [CrossRef]
- Pielou, E. Niche width and niche overlap: a method for measuring them. Ecology 1972, 53, 687–692. [Google Scholar] [CrossRef]
- Xu, Z.G.; He, Y.; Yan, B.X.; Song, C.C. Niche characteristics of typical marsh wetland plant populations in Sanjiang Plain. J. Appl. Ecol. 2007, 18, 783–787. (in Chinese). [Google Scholar]
- Wang, X.R.; Cheng, R. M.; Xiao, W. F.; Pan, L.; Zeng, L. X. Characteristics of niche changes of main dominant herbaceous plants in the initial stage of flooding in the hydro-fluctuation zones of the TGR area. Resour. Environ. Yangtze Basin, 2016; 25, 8. (in Chinese) [Google Scholar]
- Escobedo, V.M.; Rios, R.S.; Salgado-Luarte, C.; Gianoli, E. Correlation of plasticities to drought and shade: implications for environmental niche overlap in drylands. Oikos 2023, e09766. [Google Scholar] [CrossRef]
- Crain, C.M. Interactions between marsh plant species vary in direction and strength depending on environmental and consumer context. J. Ecol. 2008, 96, 166–173. [Google Scholar] [CrossRef]
- Graffmann, K.; Grosse, W.; Junk, W.J.; Parolin, P. Pressurized gas transport in Amazonian floodplain trees. Environ. Exp. Bot. 2008, 62, 371–375. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, C.; Wei, H.; Liu, Y.; Wang, Z.; Jia, Z. Effects of mixed intercropping of Cynodon dactylon and Hemarthria altissima on their biomass under different flooding conditions. Acta Ecol. Sin. 2017, 37, 54–61. [Google Scholar] [CrossRef]
- Petrů, M.; Tielbörger, K.; Belkin, R.; Sternberg, M.; Jeltsch, F. Life history variation in an annual plant under two opposing environmental constraints along an aridity gradient. Ecography 2006, 29, 66–74. [Google Scholar] [CrossRef]
- Herben, T.; Klimešová, J.; Chytrý, M. Effects of disturbance frequency and severity on plant traits: An assessment across a temperate flora. Funct. Ecol. 2018, 32, 799–808. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Ye, C.; Chen, C.; Butler, O.M.; Rashti, M.R.; Esfandbod, M.; Du, M.; Zhang, Q. Spatial and temporal dynamics of nutrients in riparian soils after nine years of operation of the Three Gorges Reservoir, China. Sci. Total Environ. 2019, 664, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.J.; Thompson, K.; Hodgson, J.G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytol. 1999, 143, 155–162. [Google Scholar] [CrossRef]
- Larbi, A.; Kchaou, H.; Gaaliche, B.; Gargouri, K.; Boulal, H.; Morales, F. Supplementary potassium and calcium improves salt tolerance in olive plants. Sci. Hortic. 2020, 260, 108912. [Google Scholar] [CrossRef]
- Nilsson, C.; Aradóttir, Á.L. Ecological and social aspects of ecological restoration: new challenges and opportunities for northern regions. Ecol. Soc. 2013, 18. [Google Scholar] [CrossRef]
- Nilsson, C.; Polvi, L.E.; Gardeström, J.; Hasselquist, E.M.; Lind, L.; Sarneel, J.M. Riparian and in-stream restoration of boreal streams and rivers: success or failure? Ecohydrology 2015, 8, 753–764. [Google Scholar] [CrossRef]
- Nilsson, C.; Svedmark, M. Basic principles and ecological consequences of changing waterregimes: riparian plant communities. Environ. Manage. 2002, 30, 468–480. [Google Scholar] [CrossRef]
- Yue, L.; Yu, C.; Abdoussalami, A.; Li, X.; Lv, K.; Huang, G.; Hu, M.; Yang, Z. Growth,Morphological Alterations, and Enhanced Photosynthetic Performance Promote Tolerance ofDistylium chinense to Alternate Drought–Flooding Stresses. Forests 2024, 15, 125. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Qin, Y.; Wang, R.; Huang, X.; Wang, Y.; Gao, T.; Yang, X. Comparative genomics reveals a unique nitrogen-carbon balance system in Asteraceae. Nat. Commun. 2023, 14(1), 4334. [Google Scholar] [CrossRef] [PubMed]
- L, D.Z.; S, Q.; Z, R.G.; W, X.P.; S, L.J.; Z, Z.L.; W, C. A. Models for niche breadth and niche overlap of species or populations. Sci. Silvae Sin. 2006, 42, 95–103. [Google Scholar]
- Li, M.X.; Wang, D.M.; Ren, Y.; Wang, X.L. Influence of different drying-rewetting frequencies on available soil nutrients and DOC. Acta Ecol. Sin. 2018, 38, 1542–1549. [Google Scholar]
- Wang, X.R.; Chen, R.M.; Xiao, W.F. ; P,L.; Zhen,L.X. Niche variation of dominant herbaceous plants in waterlevel-fluctuating zone of Three Gorges Reservoir at the beginning after charging water. Resour. Environ. Yangtze Basin, 2016; 25, 8. [Google Scholar]
- Jian, Z.J.; Ma, F.Q.; Guo, Q.S.; Qin, A.L.; Xiao, W.F. Niche of dominant plant populations in the water level fluctuation zone of canyon landform area of the Three Gorges Reservoir. Chin. J.Ecol. 2017, 36, 328–334. [Google Scholar]
- Chi, X.; Liu, J.M.; Yan, Q.; Li, P.; Ai, H.; Gao, P. Niche Characteristics of Dominant Populations of Drepanostachyum luodianense. Guizhou Agric. Sci. 2014, 42, 180–183. [Google Scholar]
- Yang, F.; Ji, D. B.; Wang, L. J.; Li, H.; Li, Y. J. Vertical distribution characteristics of dissolved oxygen and chlorophyll a in typical tributaries during the impoundment period of the Three Gorges Reservoir. Environ. Sci. 2020, 41, 2107–2115. [Google Scholar]
- Tang, T.; Cai, Q.; Liu, J. Using epilithic diatom communities to assess ecological condition of Xiangxi River system. Environ. Monit. Assess. 2006, 112, 347–361. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.L.; Wang, X.S.; Xiang, L.; Yang, J.; Min, Q.F.; Chen, G.H.; Chen, F.Q.; Huang, C.M.; Wang, G.X. Growth and respiratory metabolic adaptation strategies of riparian plant Distylium chinense to submergence by the field study and controlled experiments. Plant Physiol. Biochem. 2020, 157, 1–12. [Google Scholar] [CrossRef]
Figure 1.
Sketch map of study area and sampling sections. Notes: R1-R6 represents six transects in the RWLFZs, and N7-N9 represents three transects in the NRZs. Please refer to Table 1 for details.The lower half of the figure illustrates the hydrological variation patterns, with the left side depicting the natural hydrological fluctuation zones and the right side representing the counter-seasonally hydrological fluctuations zones..
Figure 1.
Sketch map of study area and sampling sections. Notes: R1-R6 represents six transects in the RWLFZs, and N7-N9 represents three transects in the NRZs. Please refer to Table 1 for details.The lower half of the figure illustrates the hydrological variation patterns, with the left side depicting the natural hydrological fluctuation zones and the right side representing the counter-seasonally hydrological fluctuations zones..

Figure 2.
Characteristics of soil factors at different altitudes.Different letters represent significant differences (p<0.05). SM: soil moisture, OM: organic matter, TN: total nitrogen, TP: total phosphorus, TK: total potassium, AP: available phosphorus, AK: available potassium, NO3--N: nitrate, NH4+-N: ammonium.
Figure 2.
Characteristics of soil factors at different altitudes.Different letters represent significant differences (p<0.05). SM: soil moisture, OM: organic matter, TN: total nitrogen, TP: total phosphorus, TK: total potassium, AP: available phosphorus, AK: available potassium, NO3--N: nitrate, NH4+-N: ammonium.

Figure 3.
The plant community diversity indexes in the RWLFZ and NRZ. Significance levels: NS.,P>0.05; *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. .
Figure 3.
The plant community diversity indexes in the RWLFZ and NRZ. Significance levels: NS.,P>0.05; *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. .

Figure 4.
Functional traits of vegetation in the different drawdown zones. Significance levels: NS., P>0.05; *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Pn, Net Photosynthetic Rate; SlA, Specific Leaf Area; WUE, Water Use Effciency; AGB, Above-Ground Biomass; VC,, Vegetation Coverage.
Figure 4.
Functional traits of vegetation in the different drawdown zones. Significance levels: NS., P>0.05; *, P ≤ 0.05; **, P ≤ 0.01; ***, P ≤ 0.001. Pn, Net Photosynthetic Rate; SlA, Specific Leaf Area; WUE, Water Use Effciency; AGB, Above-Ground Biomass; VC,, Vegetation Coverage.

Figure 5.
Niche overlap values for dominant herbaceous plants in the RWLFZ (A) and NRZ (B).
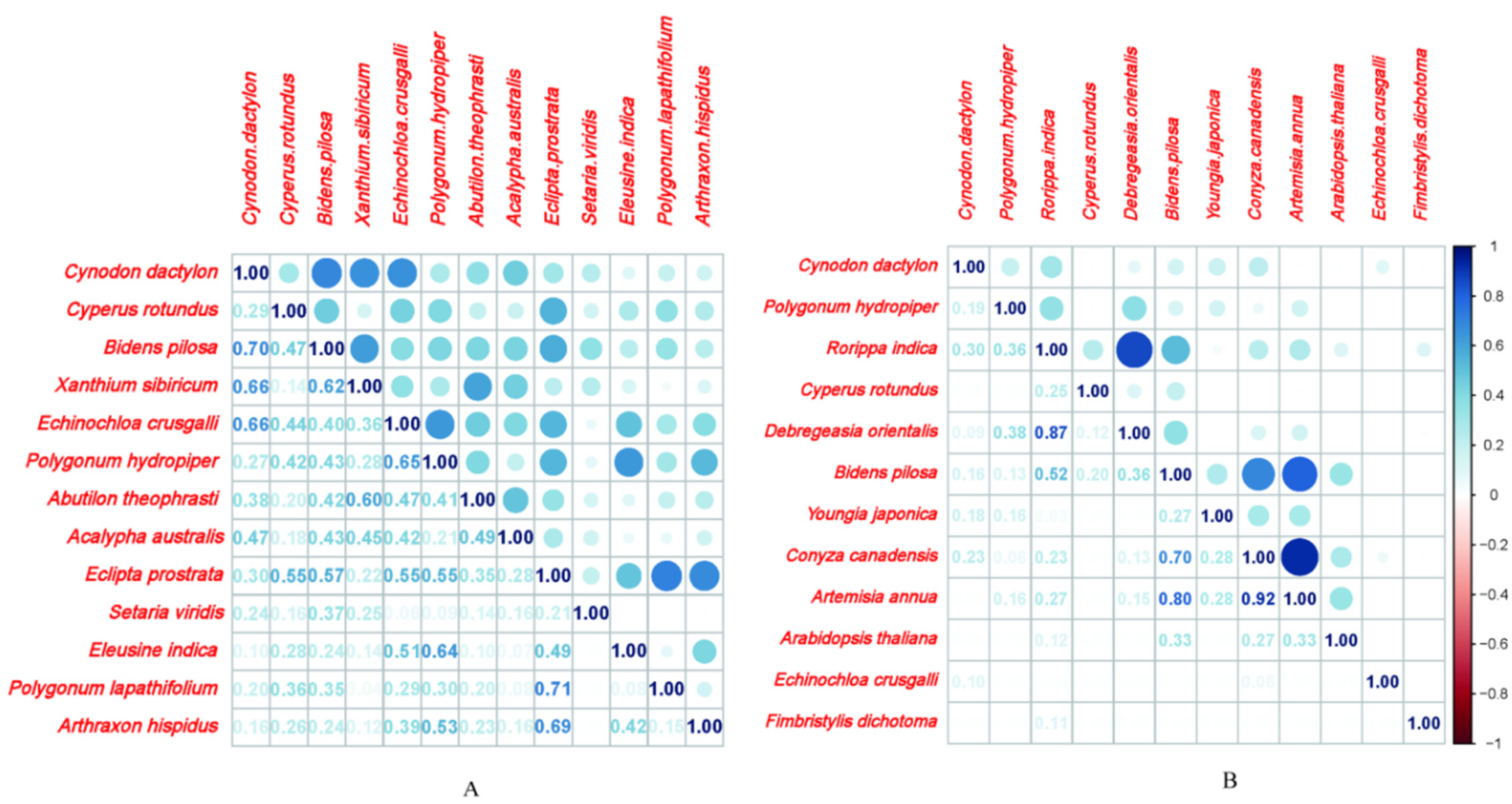
Figure 6.
DCA ordination diagram of sampling sites in the Xiangxi River (a), DCA ordination diagram of 69 species in the Xiangxi River (b).Notes: a. R1-90 sampling sites in the RWLFZ and N90-103; N91-104 sampling sites in the NRZ;b.The species represented by each number are presented in Table A2 (Appendix A).
Figure 6.
DCA ordination diagram of sampling sites in the Xiangxi River (a), DCA ordination diagram of 69 species in the Xiangxi River (b).Notes: a. R1-90 sampling sites in the RWLFZ and N90-103; N91-104 sampling sites in the NRZ;b.The species represented by each number are presented in Table A2 (Appendix A).

Figure 7.
Redundancy analysis (RDA) ordination map of the important value index of species and environment variables.The red arrow represents the environmental factor (explanatory variable), and the length of the arrow represents the intensity of the influence of the environmental factor on the community change. The longer the length, the greater the influence of the environmental factor, otherwise the opposite. The blue arrow represents the plant species (response variable). The angle between species (blue arrow) and environmental factors ( red arrow ) can represent the correlation between species and environmental factors.Environmental variables: Elevation, Flooding duration, Soil moisture, OM (organic matter), TN (total nitrogen), TP (total phosphorus), TK (total potassium), AP (available phosphorus), AK (available potassium), NO3 (nitrate), NO4 (ammonium). Species sequence in the table was the same as the order of important value of species in RWLFZ. The numbers represent the species details in Table A3 (Appendix A).
Figure 7.
Redundancy analysis (RDA) ordination map of the important value index of species and environment variables.The red arrow represents the environmental factor (explanatory variable), and the length of the arrow represents the intensity of the influence of the environmental factor on the community change. The longer the length, the greater the influence of the environmental factor, otherwise the opposite. The blue arrow represents the plant species (response variable). The angle between species (blue arrow) and environmental factors ( red arrow ) can represent the correlation between species and environmental factors.Environmental variables: Elevation, Flooding duration, Soil moisture, OM (organic matter), TN (total nitrogen), TP (total phosphorus), TK (total potassium), AP (available phosphorus), AK (available potassium), NO3 (nitrate), NO4 (ammonium). Species sequence in the table was the same as the order of important value of species in RWLFZ. The numbers represent the species details in Table A3 (Appendix A).

Table 1.
Basic characteristics of the plots.
| Sample code | Sample type | Elevation ( m ) | Slope ( ° ) | Aspect | Longitude ( ° ) | Latitude ( ° ) | Locality |
|---|---|---|---|---|---|---|---|
| R1 | RWLFZ | 145-175 | 22-27 | Northeast | 110°45'E | 30°58'N | Bazhi gate in Zigui |
| R2 | RWLFZ | 145-175 | 32-39 | Northwest | 110°45'E | 31°00'N | Wangu temple in Zigui |
| R3 | RWLFZ | 145-175 | 30-35 | East | 110°44'E | 31°02'N | Xiangjia dam in Zigui |
| R4 | RWLFZ | 170-175 | 18-29 | Northwest | 110°47'E | 31°07'N | Diaoyang rock in Zigui |
| R5 | RWLFZ | 160-175 | 16-20 | Northwest | 110°48'E | 31°07'N | Yufang flat in Zigui |
| R6 | RWLFZ | 165-175 | 10-18 | Southwest | 110°51'E | 31°11'N | Jianyangping bridge in Zigui |
| N7 | NRZ | 175-180 | 18-20 | Southwest | 110°50'E | 31°08N | Lijia mountain in Zigui |
| N8 | NRZ | 175-180 | 6-13 | Southeast | 110°49'E | 31°07'N | Shijia dam in Zigui |
| N9 | NRZ | 175-180 | 10-18 | Southwest | 110°51'E | 31°11'N | Yangdao river in Zigui |
Notes: The sample type in the table represents the type of hydrological fluctuation zone where the sampling point is located ; Elevation represents the collection of samples in that altitude range ; Slope represents the slope range of this area ; Aspect represents the orientation of the sampling point ;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.


Habitat is More Important Than Climate and Animal Richness in Shaping Latitudinal Variation in Plant Diversity
Gao Jie
et al.
,
2018
Responses of Individual Plant Species, Functional Groups, α- and β-Diversity to Short-Term Grazing Exclusion under Severe Drought Episode in Long-Term Grazed Alfa-Steppes
Yacine Kouba
et al.
,
2020
The Cape Wetland Restionaceae of New Years Peak under a Changing Climate
James Ayuk
et al.
,
2019



MDPI Initiatives
Important Links
© 2024 MDPI (Basel, Switzerland) unless otherwise stated