
Preprint
Article
Effects of Vachellia Mearnsii Tannin Extract as an Additive on Fermentation Quality, Aerobic Stability and Microbial modulation of Maize Silage
Altmetrics
Downloads
101
Views
57
Comments
0
A peer-reviewed article of this preprint also exists.


This version is not peer-reviewed
Submitted:
21 August 2023
Posted:
22 August 2023
You are already at the latest version
Alerts
Abstract
Maize silage is produced to alleviate forage shortage on dairy animals, particularly during the dry season. Microorganisms play a significant role in silage fermentation and thus determine the silage quality. Therefore, modulation of microorganisms in silage may help prohibit undesired bacteria and improve silage quality. Hence, this study evaluated the effects of tannin extract as an additive on fermentative quality, aerobic stability and microbial composition of maize silage. Therefore, a mini-silo experiment of maize with five treatments was conducted for 75 days. The results showed that additives modulate silage microorganisms. In addition, tannin extract reduced certain silage bacteria while promoting others. However, this is without affecting the silage’s fermentative quality and aerobic stability. All silage fermented well with a pH below 4.2. Tannin extract improved dry matter recovery of silage while reducing silage weight loss. Furthermore, tannin extract suppressed the establishment of undesired bacteria, such as genera Dysgonomonas, Gluconacetobacter and Clostridium, while promoting desirable bacteria, such as genera Lactobacillus and Weissella, which attributed to silage quality. Therefore, it is concluded that tannins can be strategically used as silage additives to modulate microbial composition of silage and improve silage quality by promoting the dominance of desirable bacteria in silage.

Keywords:
Subject: Biology and Life Sciences - Agricultural Science and Agronomy
1. Introduction
Over the years, ensiling has been a commonly practised technology to preserve fresh forage materials for feeding during scarcity period [1,2,3]. This is because, in silage, the loss of feeding value due to unfavourable weather conditions is very low compared to hay making [4]. According to Bernardes et al. [5], silage-making is not limited by weather conditions to such an extent that silage can be made successfully in hot or cold regions. Moreover, the benefit of ensiling is the bale technology, where storage can be accomplished without buildings [2]. Silage can be produced from different forage sources such as grasses and legumes [6], sorghum [7], maize [8], tree fodder [3,9] and agro-industrial by-products [10,11]. Generally, ensiled forage material undergoes a fermentation process where lactic acid bacteria use water-soluble sugars to produce lactic acid [12]. Lactic acid production helps rapidly reduce pH value of ensiled material to below 5 within the first three days after ensiling [4]. However, during the fermentation process, which is influenced by different microorganisms [13], silage quality may be reduced due to the degradation of protein and deamination of amino acids [14,15,16]. Subsequently, this lead to low animal protein utilisation efficiency of the ensiled forage material [17], indicating a need to modulate silage microorganisms to improve silage quality.
Likewise, the whole-maize crop is a widely used forage material for the ensiling. According to [18], maize silage has become a significant component of dairy cow ration in recent decades, and this is because of its attributes such as high biomass yield with suitable starch, good water-soluble carbohydrates and low buffering capacity [19,20]. Moreover, maize crop yield is stable under various environmental and climatic conditions and has good ensiling characteristics [18]. Generally, a whole-maize plant is suitable for ensiling because it provides adequate energy from starch in the kernel fraction and effective fibre, mainly neutral detergent fibre from the stover fraction [21]. However, the complex process that involves the interaction of forage enzymes and several microbial species during the fermentation of the ensiled forage material influences the biochemistry of the silage [19,22]. Therefore, fermentation of the ensiled material is a dynamic process which includes a series of bacterial organisms that lead to changes in silage metabolites [16,23]. Microorganisms are reported to play an essential role in the fermentation success of ensiled materials [13,21]. Hence these microorganisms determine the silage quality [24], indicating the need to study the silage's microbiology and the influence of additives at ensiling on the bacterial composition of the silage. Other studies have reported that lactic acid bacteria (LAB) inoculants affect silage microbiology by increasing the abundance of specific bacteria over others [13,25,26,27]. For example, the increase of bacteria Limosilactobacillus and Lentilactobacillus at the genus level on silages treated with LAB inoculants at ensiling has been reported [13,27,28,29]. These changes in silage microbial composition are associated with improved silage quality [25,29,30] and aerobic stability of silage after opening [13].
Therefore, manipulating silage microorganisms' compositional structure is critical in improving silage quality and nutrient use efficiency [29]. Hence, the present study evaluated the effects of tannin extract as an additive on fermentative quality (i.e. pH, dry matter recovery and silage weight loss), aerobic stability and microbial composition of whole-maize crop silage. Tannins are plant secondary metabolites that are widely distributed in plants. It is characterised by several factors, including the ability to form complexes with protein (bind protein) and polysaccharides due to hydroxyl phenolic groups present [31,32,33]. In addition to that, tannins are antimicrobial, anti-parasitic, and it has immunomodulatory properties [34]. The use of tannin extract as a silage additive has been reported on bacterial community structure and proteolysis in alfalfa silage [16]. Jayanegara et al. [9] concluded that tannins have the potential to be used as silage additives during ensiling to reduce proteolysis during the fermentative stage. But little has been reported on the effects of tannins on the fermentative quality, and bacterial modulation of high energy forage material. Therefore, in this study, Vachellia mearnsii (formerly known as Acacia mearnsii) tannin extract was used as an additive to maize silage to evaluate its ability to modulate silage microbial composition and the contribution of that toward the silage quality. Hence, it was hypothesized that tannin extract could effectively modulate the microbial composition of whole-maize crop silage without affecting fermentation quality and aerobic stability.
2. Materials and Methods
2.1. Study Area and Ethics Approval
The study was conducted at the Agricultural Research Council-Animal Production (ARC-AP), Irene farm in South Africa. The details of the study's geographical location and climatic conditions are provided by Mpanza et al. [35]. The ethics review, and approval were waived because the study did not include the use of animals.
2.2. Maize Harvesting, Treatment Preparation and Ensiling
Zea mays are planted yearly at ARC-AP farm in Irene, South Africa, mainly to produce silage to feed our dairy cows during the dry season. Maize silage helps ensure that there is high-energy feed for animals during the dry season. Therefore, for this study, maize used for silage was planted in November 2021 at ARC-AP farm, and the whole-maize crop was harvested at the half-milk growth stage (36.9% DM) in mid-February 2022. Feraboli 945 forage harvester (Fondada Nel, Cremona, Italy) was used to harvest and chop the forage to a theoretical length of 2 -3 cm. The chopped maize forage was divided into five equal portions of 4 kg each on a fresh mass basis. Each group was assigned to one of the five treatments, which were maize forage alone without inoculant (i.e. LAB or tannin extract), referred as negative control (T1), maize forage inoculated with LAB plus 1% tannin extract (T2), maize forage inoculated with LAB referred as positive control (T3), maize forage inoculated with LAB plus 2% tannin extract (T4) and maize forage inoculated with LAB plus 3% tannin extract (T5). The tannin extract inclusion percentage was calculated per kilogram of maize forage on fresh matter bases. Tannin extract powder was spread on chopped maize forage and thoroughly mixed by hand with the maize forage and immediately ensiled.
The tannin extract used in this study was donated by UCL (Pty) Ltd Company, based in Dalton in Kwa-Zulu Natal, South Africa. Tannin was extracted from Vachellia mearnsii (Black wattle) barks by UCL Company, and it is usually referred to as condensed tannin or Mimosa extract. The tannin extract contained about 66% condensed tannin on a dry matter basis. Therefore, the 1%, 2% and 3% of tannin inclusion levels on maize silage are equivalent to 6.6 g, 13.2 g and 19.8 g of condensed tannin inclusion per kg of fresh maize forage, respectively. On the other hand, the lactic bacteria inoculant that was used during ensiling was Kem LAC HD Dry (Kemin Industries, South Asia (Pty) Ltd), which hereafter will be referred to as lactic acid bacteria (LAB). The Kem LAC HD Dry is a white powder containing lactic acid-producing bacteria, including Lactobacillus plantarum, Lactobacillus acidophilus and Lactobacillus bulgaricus. All these bacteria have the same concentration of 1.36 x109 Colony Forming Units per gram (CFU/g). The LAB inoculant is reported to help improve silage fermentative quality by speeding up lactic acid fermentation, reducing proteolysis, controlling silage temperature, and enhancing aerobic stability and DM retention [36]. The Kem LAC HD Dry inoculant was purchased from OBARO (Pty) Ltd. Pretoria in South Africa. The amount of LAB used as an inoculant in this study was based on a company recommendation of 1.5 g per ton of silage on a fresh mass basis. The LAB inoculant was dissolved in 20 mL of distilled water on the same day of ensiling. The negative control was sprayed with only 20 mL of distilled water to standardize silage moisture content. Approximately 970 g of maize forage was ensiled in a pre-weighed 1.5 L glass jar each, with each treatment replicated 3 times. The ensiled material was stored at room temperature (23-25 °C) for 75 days.
2.3. Characterization of Fresh Forage and Silage
On day zero (ensiling day), about 80 g of fresh samples (three replicates per treatment) were taken. The 80 g samples were divided into two equal portions; one portion (40 g) was mixed with 350 mL distilled water, vigorously shaken for 2 minutes, and kept in a fridge at 5 °C overnight. The mixture was filtered through a Whatman No 1 filter paper the following day. The extract was used to determine pH with a pH meter (Thermo Orion Model 525, Thermo Fisher Scientific Waltham. MA, USA). Another 40 g portion was oven dried at 60 °C for 72 hours to determine dry matter content at ensiling.
After 75 days of ensiling, all bottle jars with silage were weighed to estimate the weight loss per treatment due to fermentation. Three bottle jars of the same treatment were opened, the top 2 cm spoiled silage was removed and discarded, the rest was emptied into a 10 L bucket at a time, and thoroughly hand mixed. To prevent cross-contamination, plastic hand gloves were used each time we opened the silage treatment and discarded afterwards. Silage pH and dry matter were determined, as explained above. To assess the aerobic stability of silage, an extra 100 g of fresh silage (three replicates per treatment) was put loosely in a cleaned plastic container and exposed to air for four days. A thermometer was inserted into the geometric centre of the silage to record the silage temperature. It was covered with nylon to prevent dust contamination while allowing airflow, as described by [37]. Containers with silage were kept at room temperature of 24 °C. Silage would not be considered aerobically stable when the silage temperature rises 2 °C above room temperature within four days of air exposure. The time taken (in hours) for silage temperature to exceed room temperature by 2 ºC would be recorded. In addition, about 15 g (in duplicates per treatment) of silage sample was taken into a plastic container closed tightly and frozen at –80 °C until used for DNA extract (see section 2.4 below for details). Dry matter recovery was determined by calculating the quotient between the weight of the silage at day 75 and the weight of the forage at day 0, corrected for their respective dry matter content. While weight losses due to fermentation were determined by calculating the difference between silage weights at day 75 and the weight of fresh forage at day 0, corrected for dry matter content, respectively.
2.4. Deoxyribonucleic Acid (DNA) Extraction and Sequencing Analysis
Microbial DNA was extracted with Macherey-Nagel™ NucleoSpin™ DNA Stool kit following the manufacturer's guidelines, and DNA concentration was evaluated with Nanodrop 2000 (Thermo Electron Corporation, USA). The 16S Metagenomics Sequencing Library Preparation guide (Illumina, San Diego, USA) was followed to perform 16S rRNA amplification and sequencing. The V3-V4 hypervariable regions were amplified using primers with adapters (Forward= 5'TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG3', Reverse= 5'GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC 3'). The PCR was conducted using the reaction procedures detailed by Mpanza et al. [35]. The purified PCR products were sequenced using the Illumina MiSeq platform and generated 300 bp paired-end reads. The raw reads were trimmed to remove low-quality sequences with Trimmomatic version 0.36. QIIME2 software was employed for downstream analysis, pre-processing paired-end sequence reads using the DADA2 pipeline, including QC, denoising, merging sequences, and removing chimeric sequences. Sequences were clustered de novo, and sequences with ≥ 97% similarity were assigned to the same operational taxonomic units (OTUs). The OTU feature table was generated, and representative OTU sequences were aligned to the Greengenes database for taxonomic assignment.
2.5. Statistical Data Analysis
Data on pH, dry matter recovery, weight loss and aerobic stability were subjected to one-way analysis of variance (ANOVA) using SAS version 9.0 (SAS Institute, Inc., Cary, NC, USA). The following statistical model was used:
where Yij is a general observation, µ is a general mean, ġi is the additives effects (i= LAB, tannin extract), and εij is the random error. Regarding a significant difference between treatment means, the PDIFF statement was used to compare the means. Significant was declared at p ≤ 0.05, and the tendency at 0.05 < p ≤ 0.1.
Yij = µ + ġi + εij
Bioinformatics data analysis on the microbial composition of silage was done using RStudio with R core (version 4.2.2). Alpha diversity matrices (Observed and Shannon) were calculated and tested for variances using the Kruskal-Wallis test. Beta diversity was calculated using distance matrices generated from unweighted UniFrac analysis.
3. Results
3.1. Fermentative Charactereistics of Maize Silage
The fermentative characteristics of maize silage were measured by determining silage pH, dry matter (DM), Dry matter recovery (DMR) and weight loss (WL) on opening day (i.e. day 75). After opening, the silage was exposed to air to determine aerobic stability. Table 1 shows the fermentation quality of maize silage treated with different additives. These results showed that the pH value at ensiling (day 0) was above 5, as was expected since no fermentation had occurred. It was noticed that treatment 5 had a significantly (p < 0.05) higher pH value compared to the other treatments. However, it is worth noting that the pH of all silage treatments dropped to below 4.0 at day 75 after ensiling. Therefore, tannin extract as an additive did not affect the fermentative characteristics of maize silage. However, it was observed that there was a significant (p < 0.05) variation in pH values among the treatments at day 75, with treatment 5 recorded as the highest.
Furthermore, this study showed a significant (p < 0.05) variation in silage dry matter content. Treatments 4 and 5, treated with 2% and 3% of tannin extract, had high DM. It was observed that the tannin extract additive did not affect dry matter recovery (DMR) of the silage, however there was a tendency to improve DMR with a p-value of 0.0624. Although treatments 2 and 4, silage recorded a 13.1% and 14.0% higher DMR, respectively, as compared to silage in treatment 1 (negative control) but it was not statistically significant (p > 0.05). A similar pattern was observed with silages in treatments 3 and 5 recorded a numerically higher DMR than the negative control, but it was not statistically significant (p > 0.05). A similar pattern was also observed in weight loss (WL) due to fermentation. Tannin extract as an additive did not affect WL of maize silage, even though treatments 2 and 4, silages reduced weight loss by 68.7% and 67.2%, respectively, compared to the negative control treatment (maize silage without additives).
Figure 1 shows the temperature of maize silage exposed to air for four days after opening. Silage treated with LAB (positive control) or LAB plus tannin extract did not affect silage aerobic stability within the four days of silage exposure to air. None of the silage treatments recorded a temperature above the room temperature of 24 °C within the four days of exposure to air.
3.2. Silage Microbial Composition as Influenced by Additives
A total of 46 OTUs were obtained from the samples. The abundance of the bacterial composition was estimated across treatments, with 3 phyla detected at a relative abundance of ≥ 0.05% (Figure 2a). Lactic acid bacterial inoculation improved the abundance of the Bacteriodetes phylum while reducing the Proteobacteria phylum (positive control treatment 3). However, what was noticeable was that the tannin extract additive improved the relative bacterial abundance of phylum Firmicutes on maize silage in T2, T4 and T5. The phylum Firmicutes abundance level recorded for T1, T2, T4 and T5 were 38.8%, 97.2%, 83.7% and 85.1%, respectively. This indicates an increase of 2.2 to 2.5 times the value recorded for Firmicutes in T1 (negative control) silage. On the other hand, treatment 3 (positive control) showed a different pattern where phylum Bacteriodetes was predominant, accounting for 50.7%, and Firmicutes phylum was the second most abundant with 41.7%. Bacteriodetes phylum in other silage treatments (T1, T2, T4 and T5) were 34%, 1.2%, 14.2% and 12.2%, respectively. Proteobacteria accounted for 27.2%, 3%, 15.2%, 2.1% and 2.8% in T1, T2, T3, T4 and T5, respectively.
At the genus level, 13 genera were identified (Figure 2b). This study shows a shift in bacteria abundance at the genus level associated with the silage additive. Inoculating maize silage (T3 positive control) with LAB improved the abundance of genera Dysgonomonas and Weissella compared with maize silage in T1 (negative control). This study recorded a relative abundance of 29.1% and 47.3% for Dysgonomonas and 3.5% and 7.9% for Weissella in treatments 1 and 3, respectively. This indicates an increase of 1.6 and 2.3 times the values recorded for these genera in T1 silage. On the other hand, silage that was treated with tannin extract at ensiling favoured the dominancy of the Lactobacillus genus accounting for 63.7%, 63.5% and 73.2% in T2, T4 and T5 silages. This indicates an increase of more than double the value recorded in T1 (negative control) silage (30.2%). Similarly, the genus Weissella increased the value recorded for silage in treatment 1 by six times. Silage additives affected genus Clostridium, as LAB reduced the abundance by 2.5 times, whereas it was not detected in silage treated with tannin extract. Other genera such as Pediococcus (5.1%, 2.6% and 3%) and Leuconostoc (1.9%, 2.6% and 0.9%) were recorded on silage treated with tannin extract (i.e. T2, T4 and T5).
There was no significant (p > 0.05) difference in the alpha diversity of the bacterial community in different silage treatments (data not shown). Figure 3 shows the principal coordinate analysis (PCoA) of bacterial composition on maize silage as influenced by additives. The PCoA showed that axis 1 accounted for 38.6% variation of bacterial differences due to negative control treatment (T1 silage), whereas bacterial differences between T2, T3, T4 and T5 silages accounted for 24.6% variation of axis 2. The PCoA portrays a clear cluster separation of bacterial composition observed between treatments. However, there was an overlap in the clustering of bacteria for treatments 2 and 3 (positive control). This study showed that treatments 4 and 5 bacteria were clustered separately from others.
The bacterial composition's canonical correspondence analysis (CCA) in maize silages is depicted in Figure 4, showing a clear variation in silage bacterial composition between the treatments. The CCA 1 showed that 37.1% of the bacterial variation between treatments was due to negative control treatment (T1), whereas CCA 2 showed that T3 (positive control) was responsible for 22.4% of the bacterial variation. The CCA plot showed no relationship between T1 and T3 bacteria, and bacteria in maize silage treated with tannin extract (i.e. T2, T4 and T5) had a strong relationship as they were clustered together.
The analysis of similarities between the bacterial compositions of silage treatments as influenced by additives is presented in Figure 5. The ANOSIM showed a significant (p < 0.05) dissimilarity between bacterial compositions of maize silages subjected to different treatments.
Figure 6 shows common and unique operation taxonomic units (OTUs) between maize silages of different treatments. Venn diagram showed that there were only 4 OTUs that were shared among all the silage treatments, while most OTUs were unique to each treatment. However, silage in treatment 2 showed only 2 unique OTUs. Tannin-treated silages shared 3 OTUs, while treatments 4 and 5 shared 2 OTUs.
4. Discussion
Generally, the pH value of the silage is one of the parameters used to judge the fermentative quality and an indicator of a well-preserved silage. A well-fermented silage recommends having a pH value of 4.2 or below [19]. This is because, during fermentation, organic acids (mainly lactic acid) are produced by lactic acid bacteria, which helps in a drastic reduction of pH to below 5 within the first three days of ensiling [4,12,38]. In this study, the pH values at day zero (ensiling day) were above 5; at day 75, the post-ensiling pH values were below 4.2 (Table 1). Therefore, in this study, all treatments produced sufficient lactic acid required to reduce pH, indicating well-fermented silage with good fermentative quality. The final pH recorded on different silage treatments in this study was in the range of an adequately fermented silage below 4.2 [28]. In addition, a pH value below 4.2 helps reduce the chances of unwanted microorganisms in silage [39,40]. Therefore, it is safe to say that Vachellia mearnsii tannin extract did not interfere with silage fermentation characteristics of maize silage; hence maize silage treated with tannin extract fermented well. This is because silage pH is considered the critical indicator of the fermentation quality of the ensiled forage material [3] and for an adequately preserved silage [41]. In addition, Vachellia mearnsii tannin extracts additives had no effects on aerobic stability of the silage.
The dry matter (DM) content of maize at ensiling was ranging from 34.7% to 40.2% but within the range of 28% to 40% recommended for forage at ensiling [42]. On the opening day (day 75), the silage had a DM content ranging from 31.1% to 36.5%, above 28%, the minimum recommended DM for silage [42,43]. Therefore, this indicates that all maize silages were preserved well [42]. However, maize silage from treatments 4 and 5 had a significantly higher DM percentage than other treatments (Table 1). The present study showed that treating maize at ensiling with tannin extract had no significant effect on dry matter recovery and weight loss of silage due to fermentation. This is despite the fact that tannin treated silage showed numerically higher DMR and lower WL. Silage dry mater recovery coupled with low weight loss indicate low moisture content and such silage is reported to maintain a low temperature, especially during aerobic exposure [44]. This agrees with our results for aerobic stability; all treatments recorded temperatures below the ambient temperature over four days of aerobic exposure (Figure 1). However, this results must be interpreted with caution due to the fact that silage was exposed to air for four days and we don’t know how long the silage would have maintained aerobic stability if were exposed longer than four days. On the other hand, reducing weight loss of the ensiled forage materials is regarded as a biological benefit, for it improves silage production [7]. Therefore, in this study, tannin inclusion levels of 1% and 2% showed a tendency to improve silage production by reducing the weight loss of maize silage.
Silage fermentative characteristics (i.e. pH, short-chain fatty acids, DMR and weight loss) are good indicators of well-preserved silage; however, this does not necessarily determines the quality of silage. Therefore, it is essential to characterize what is in the silage to determine the quality. On the other hand, microbes have been reported as drivers of silage quality depending on the substrates present during ensiling [21,24]. Therefore, molecular characterization of silage microorganisms is essential to know the microbial composition of silage, as the dominant bacteria in silage has been reported to determine the silage's quality [24,45]. Therefore, genomics technology offers an opportunity to study the microbiological aspects of the ensiled forage material to determine the silage's microbial composition. Subsequently, in this study, a 16S ribosomal RNA gene sequencing was conducted in different maize silage treatments ensiled for 75 days to evaluate the effects of treatments on microbial composition.
Microbial characterization of maize silage revealed that silage additives influenced the bacterial composition of the silage (see Figure 4, Figure 5 and Figure 6). The high relative abundance of a few dominant phyla (Bacteroidetes, Firmicutes and Proteobacteria) recorded in this study was attributed to a decrease in bacterial composition, which allied with the additives used during ensiling. Results reported in this study showed that silage additives led to a shift in bacterial dominance between the treatments. Applying LAB inoculant (treatment 3) on maize silage at ensiling favoured the dominancy of phylum Bacteroidetes bacteria followed by phyla Firmicutes and Proteobacteria. On the contrary, Jaipolsaen et al. [46] and Dong et al. [47] reported Firmicutes bacteria as the dominant phylum on maize silage treated with LAB inoculant at ensiling. However, other studies reported Proteobacteria as the dominant phylum on silage produced from different forages treated with LAB inoculant at ensiling [13,21,39]. The variation in bacterial dominance reported by various studies can be attributed to the LAB substrates used as an additive and the forage material used for silage production. The current study further showed that including tannin extract as a silage additive affected the microbial composition of silage, favouring the dominance of phylum Firmicutes. These results agree with other studies that reported the dominancy of phylum Firmicutes on silage produced from different forage materials [39,48,49,50,51,52]. Phylum Firmicutes constituted 84% to 97% of the bacterium detected on maize silage in treatments 2, 4 and 5. While in treatments 1 and 3 maize silage, phylum Firmicutes constituted 39% and 42% of the detected bacteria. The dominance of Firmicutes in silage was reported as an indicator of well-fermented and good-quality silage [53], and this is because Firmicutes are reported to have the potential to secrete various enzymes under anaerobic conditions [53], which are involved in degrading the lignin content of the ensiled forage [54,55].
Low pH in silage indicates well-preserved silage and prevents undesired bacterial growth [52]; this was observed in the current study as all treatments recorded pH below 4.0. However, microbial characterisation of these silages using genomic technology analysis revealed the presence of undesirable bacteria in T1 (negative control) and T3 (positive control) silages. Thus, this study detected the bacteria Dysgonomonas, Gluconacetobacter and Clostridium, classified as undesirable genera. In the T1 silage, these bacteria accounted for about 65% of the total detected genera. In T3 silage, genera Dysgonomonas and Clostridium were detected, accounting for about 50% of the detected bacteria. The presence of these undesirable genera in silage compromises silage quality despite its fermentative quality [39,56]. Dysgonomonas genus was reported to occur in response to circumstances rather than naturally [56]; therefore, it is necessary to study its presence in silage. Genus Gluconacetobacter is reported to favour acetic acid production, which increases silage pH [39]. Clostridium bacteria are reported to be involved in the production of alcohol in silage [57]. Furthermore, Clostridium bacteria decompose protein into ammonia nitrogen and lead to protein loss from the silage [58]. Therefore, the presence of Clostridium bacteria in silage is associated with several problems when such silage is fed to animals; these include nitrogen pollution due to high ammonia [59], ketonemia and reduction of milk production in cattle [60].
On the other hand, tannin-treated silages were dominated by the genus Lactobacillus bacteria, which is associated with well-preserved silage [40]. Lactobacillus is recorded as an essential microorganism during ensiling due to its ability to control lactic fermentation [39,61]. Therefore, in high-quality silage, Lactobacillus is the predominant bacteria [62]. Likewise, in this study, Lactobacillus was abundant in tannin-treated silage treatments. Weissella bacteria was the second dominant genus in maize silage treated with tannin extract, accounting for 31.3%, 22.8% and 21.1% in treatments 2, 4 and 5, respectively. Lactobacillus and Weissella bacteria are reported to influence lactic fermentation by producing lactic acid during ensiling [27]. Hence, these two genera are reported to contribute significantly to reducing silage pH, particularly in the early phase of silage [30,48]. In addition, Weissella bacteria are reported to convert soluble carbohydrates into carbon dioxide and water during silage fermentation [40]. The dominance of Lactobacillus and Weissella genera reported in this study on maize silage treated with tannin extract concurs with the result reported by [58].
The analysis of PCoA and CCA showed that additives, either LAB or tannin extract, plays a critical role in the bacterial community structure of the silage. The CCA clearly showed that treating maize silage with tannin extract at ensiling favours a particular type of bacteria (i.e. Lactobacillus and Weissella from phylum Firmicutes) which are critical for silage quality. ANOSIM also showed significant (p < 0.006) dissimilarities between the bacterial structure of maize silage as influenced by additives. Venn diagram also showed uniqueness in the OTUs as influenced by silage treatments (additives). A well-preserved silage was reported to be dominated by phylum Firmicutes and genera Lactobacillus and Weissella bacteria [30]. Therefore, a similar pattern has been observed in the present study, where maize silage treated with tannin extract improves silage quality by modulating silage microorganisms.
5. Conclusions
This study aimed to determine the effects of Vachellia mearnsii tannins extract additive on fermentative quality, aerobic stability and microbial composition of maize silage. All silage fermented recorded a pH below 4.0 at day 75 post-ensiling. This study showed that tannin extract could be strategically used as a silage additive to improve silage quality without affecting the fermentative quality of the silage. Adding tannin extract in maize silage at ensiling prohibited the growth of undesirable bacteria while promoting the growth of desired bacteria. The dominancy of genera Lactobacillus and Weissella bacteria in maize silage treated with tannin extract is good evidence of the feasible strategy in modulating silage bacterial composition while improving silage quality. Further study is required to determine the effects of feeding such silage on animal feed intake, digestibility, rumen modulation, methane emission, blood metabolites and growth performance.
Author Contributions
Conceptualization, T.D.E.M.; methodology, T.D.E.M.; software, S.M.; validation, S.M. and T.D.E.M; formal analysis, S.M. and T.D.E.M.; investigation, T.D.E.M.; resources, T.D.E.M; data curation, S.M.; writing—original draft preparation, T.D.EM and S.M.; writing—review and editing, T.D.E.M. and S.M.; visualization, S.M.; funding acquisition, T.D.E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Department of Agriculture, Land Reform & Rural Development (DALRRD), Republic of South Africa, the project number P02000233.
Data Availability Statement
The sequences presented in this study were deposited to the National Center of Biotechnology Information (NCBI) Sequence Read Archive (SRA) database under Bioproject PRJNA976900, accessible at this link: https://www.ncbi.nlm.nih.gov/sra/PRJNA976900.
Acknowledgments
Authors would like to acknowledge the kind support of the donation of tannin extract from UCL (Pty) Ltd, a Company, based in Dalton in Kwa-Zulu Natal, South Africa.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, Y.; Zhao, X.; Chen, W.; Zhou, Z.; Meng, Q.; Wu, H. Effects of adding various silage additives to whole corn crops at ensiling on performance, rumen fermentation, and serum physiological characteristics of growing-finishing cattle. Animals, 9. [CrossRef]
- Jayanegara, A.; Sujarnoko, T.U.P.; Ridla, M.; Kondo, M.; Kreuzer, M. Silage quality as influenced by concentration and type of tannins present in the material ensiled: A meta-analysis. J. Anim. Physiol. Anim. Nutr. [CrossRef]
- Wang, C.; Pian, R.; Chen, X.; Lv, H.; Zhou, W.; Zhang, Q. Beneficial effects of tannic acid on the quality of bacterial communities present in high-moisture Mulberry leaf and Stylo silage. Front. Microbiol. 2020, 6412. [Google Scholar] [CrossRef] [PubMed]
- Ntakyo, P.R.; Kirunda, H.; Tugume, G.; Natuha, S. Dry Season Feeding Technologies: Assessing the Nutritional and Economic Benefits of Feeding Hay and Silage to Dairy Cattle in South-Western Uganda. Open J. Anim. Sci. 2020, 10, 627–648. [Google Scholar] [CrossRef]
- Bernardes, T.F.; Daniel, J.L.P.; Adesogan, A.T.; McAllister, T.A.; Drouin, P.; Nussio, L.G.; Huhtanen, P.; Tremblay, G.F.; Bélanger, G.; Cai, Y. Silage review: Unique challenges of silages made in hot and cold regions. J. Dairy Sci, 4: 101, 4001. [Google Scholar] [CrossRef]
- Kaewpila, C.; Khota, W.; Gunun, P.; Kesorn, P.; Kimprasit, T.; Sarnklong, C.; Cherdthong, A. Characterization of green manure sun hemp crop silage prepared with additives: aerobic stability, nitrogen value and in vitro rumen methane production. Fermentation. 2022, 2022. 8, 104. [Google Scholar] [CrossRef]
- Sun, Z.; Li, Y.; Liu, G.; Gao, R.; Bao, J.; Wang, L.; Wu, Z.; Yu, Z. Associative effects of ensiling mixtures of sweet sorghum and korshinsk pea shrub on fermentation quality, chemical composition and in vitro rumen digestion characteristics. Anim. Sci. J, e: 93, 1370. [Google Scholar] [CrossRef]
- Guan, H.; Shuai, Y.; Yan, Y.; Ra, Q.; Wang, X.; Li, D.; Cai, Y.; Zhang, X. Microbial community and fermentation dynamics of corn silage prepared with heat-resistant lactic acid bacteria in a hot environment. Microorganisms. 2020, 2020. 8, 719. [Google Scholar] [CrossRef]
- Jayanegara, A.; Yaman, A.; Khotijah, L. Reduction of proteolysis in high protein silage from Maringa and Indigofera leaves by addition of tannin extract. Vet. World. [CrossRef]
- Sadarman, S.; Ridla, M.; Nahrowi, N.; Ridwan, R.; Jayanegara, A. Evaluation of ensiled soy source by-product combined with several additives as an animals feed. Vet. World. 2020, 2020. 13, 940–946. [Google Scholar] [CrossRef]
- Schneider, C.R.; Zambom, M.A.; Galhardo, D.; Faccenda, A.; Avila, A.S.; Tinini, R.C.R.; Del Valle, T.A. Microbiological quality of silage made from by-products of cassava starch extraction and viticulture. S. Afr. J. Anim. Sci. 2021, 51(3), 407–415. [Google Scholar] [CrossRef]
- Kung Jr, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical microbial, and organoleptic components of silages. J. Dairy Sci, 4: 101, 4020. [Google Scholar] [CrossRef]
- Tahir, M.; Li, J.; Xin, Y.; Wang, T.; Chen, C.; Zhong, Y.; Zhang, L.; Liu, H.; He, Y.; Wen, X.; Yan, Y. Response of fermentation quality and microbial community of oat silage to homofermentative lactic acid bacteria inoculation. Front. Microbiol, 1: 13, 1091. [Google Scholar] [CrossRef]
- Ke, W.C.; Ding, W.R.; Xu, D.M.; Ding, L.M.; Zhang, P.; Li, F.D.; Guo, X.S. Effects of addition of malic or citric acids on fermentation quality and chemical characteristics of alfalfa silage. J. Dairy Sci. 2017, 100(11), 8958–8966. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Jiang, Y.; Kim, D.H.; Pech Cervantes, A.A.; Arriola, K.G.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C. ; Adesogan. A.T. Fate of Escherichia coli O157:H7 and bacterial diversity in corn silage contaminated with the pathogen and treated with chemical or microbial additives. J. Dairy Sci. 2017. 100, 1780-1794. [CrossRef]
- Ke, W.; Zhang, H.; Li, S.; Xue, Y.; Wang, Y.; Dong, W.; Cai, Y.; Zhang, G. Influence of Condensed and Hydrolysable tannins on the bacterial community, protein degradation and fermentation quality of alfalfa silage. Animals. 2022, 2022. 12, 831. [Google Scholar] [CrossRef]
- Getachew, G. Depeters, E.J.; Pittroff, W.; Putnam, D.H.; Dandekar, A.M. Review: Does protein in alfalfa need protected from rumen microbes? Prof. Anim. Sci. [CrossRef]
- Khan, N.A.; Yu, P.; Ali, M.; Cone, J.W.; Hendriks, W.H. Review: Nutritive value of maize silage in relation to dairy cow performance and milk quality. J. Sci. Food Agric. [CrossRef]
- McDonald, P. , Henderson A.R., and Heron S.J.E. The biochemistry of silage. 1991. 2nd ed. Welton, Lincoln, UK: Chalcombe Publication.
- Silva, L.D.; Pereira, O.G.; Silva, T.C.; Leandro, E.S.; Paula, R.A.; Santos, S.A.; Ribeiro, K.G.; Valadares-Filho, S.C. Effects of Lactobacillus buchneri isolated from tropical maize silage. Grass Forage Sci. [CrossRef]
- Li, H.; Zeng, T.; Du, Z.; Dong, X.; Xin, Y.; Wu, Y.; Huang, L.; Liu, L.; Kang, B.; Jiang, D.; Wu, B.; Yang, W.; Yan, Y. Assessment on the Fermentation Quality and Bacterial Community of mixed Silage of Faba Bean with Forage Wheat or Oat. Front. Microbiol, /: 13, 875819. https, 8758; 13. [Google Scholar]
- Xin, Y. F.; Chen, C.; Zeng, T. R.; Du, Z. C.; Ni, H. R.; Zhang, Y. H.; et al. Research progress in effects of additives to silage on microbial diversity. Biotechnol. Bull. 2021, 37, 24–30. [Google Scholar] [CrossRef]
- Xu, D.; Ding, W.; Ke, W.; Li, F.; Guo, X. Modulation of metabolome and bacterial community in whole crop corn silage by inoculating homofermentative Lactobacillus plantarum and Heterofermentative Lactobacillus buchneri. Front. Microbiol. [CrossRef]
- Ridwan, R.; Abdelbagi, M.; Sofyan, A.; Fidriyanto, R.; Astuti, W.D.; Fitri, A.; Sholikin, M.M.; Rohmatussolihat Sarwono, K.A.; Jayanegara, A.; Widyastukt, Y. A metta-analysis to observe silage microbiome differentiated by the use of inoculant and type of raw material. Front. Microbiol. 2023. [Google Scholar] [CrossRef]
- Yan, Y.; Li, X.; Guan, H.; Huang, L.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.; Yang, W.; Cai, Y.; Zhang, X. Microbial community and fermentation characteristics of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Wang, X.; Lin, Y.; Yang, X.; Ni, K.; Yang, F. Microorganisms that are critical for the fermentation quality of paper mulberry silage. Food energy Secur. 2021. 10, e304. [CrossRef]
- Xian, Z.; Wu, J.; Deng, M.; Wang, M.; Tia, H.; Liu, D.; Li, Y.; Liu, G.; Sun, B.; Guo, Y. Effects of cellulase and Lactiplantibacillus plantarum on the fermentation parameters, nutrients, and bacterial community in Cassia alata silage. Front. Microbiol. 2022, 13, 926065. [Google Scholar] [CrossRef]
- Zhao, S.; Yang, F.; Wang, Y.; Fan, X.; Feng, C.; Wang, Y. Dynamics of fermentation parameters and bacterial community in high-moisture alfalfa silage with or without lactic acid bacteria. Microorganisms. 2021, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lui, Y.; Wang, Z.; Bao, J.; Zhao, M.; Si, Q.; Sun, P.; Ge, G.; Jia, Y. Effects of Different Types of LAB on Dynamic Fermentation Quality and Microbial Community of Native Grass Silage during Anaerobic Fermentation and Aerobic Exposure. Microorganisms. 2023, 11, 513. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Sun, L.; Wang, Z.; Wang, W.; Xin, X.; Xu, L.; Du, S. Fermentation Characteristics, Microbial Compositions and Predicted Functional Profiles of Forage Oats Ensiled with Lactiplantibacillus plantarum or lantilactobillus buchneri. Fermentation. 2022, 8, 707. [Google Scholar] [CrossRef]
- Naumann, H.D.; Tedeschi, L.O.; Zeller, W.E.; Huntley, N.F. The role of condensed tannins in ruminant animal production: advances, limitations and future directions. R. Bras. Zootec, 46. [CrossRef]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr, 1: 4. [CrossRef]
- Fonseca, N.V.B.; Cardoso, A.S.; Bahia, A.S.R.S.; Messana, J.D.; Vicente, E.F.; Reis, R.A. Additive tannins in ruminant nutrition: An alternative to achieve sustainability in animal production. Sustainability 2023, 15, 4162. [Google Scholar] [CrossRef]
- Maisetta, G.; Batoni, G.; Caboni, P.; Esin, S.; Rinaldi, A.C.; Zucca, P. Tannin profile, antimicrobial properties, and antimicrobial activity of extracts from two Mediterranean species of parasitic plant Cytinus. BMC Complement. Altern. Med. 2019, 2019. 19, 82. [Google Scholar] [CrossRef]
- Mpanza, T.D.E.; Dhlamini, T.C.; Pierneef, R.E.; Mbatha, K.R. Enteric methane emission, rumen fermentation and microbial profiles of meat-master lambs supplemented with barley fodder sprouts. Fermentation, 8. [CrossRef]
- Aragón, Y.A.; Jatkauskas, J.; Vrotnaikienė, V. The effect of a silage inoculant on silage quality, aerobic stability and meat production on farm scale. ISRN. Vet. Sci. 2012, 5927. [Google Scholar] [CrossRef]
- Li, F.; Ding, Z.; Adesogan, A.T.; Ke, W.; Jiang, Y.; Bai, J.; Mudassar, S.; Zhang, Y.; Huang, W.; Guo, X. Effects of class 11a bacteriocin-producing Lactobacillus species on fermentation quality and aerobic stability of alfalfa silage. Animals. [CrossRef]
- Liu, Q.; Li, X.; Desta, S.T.; Zhang, J.; Shao, T. Effects of Lactobacillus plantarum and fibrolytic enzyme on the fermentation quality and in vitro digestibility of total mixed ration silage including rape straw. J. Integr. Agric. 2016, 2016. 15, 2087–2096. [Google Scholar] [CrossRef]
- Zi, X.; Li, M.; Chen, Y.; Lv, R.; Zhou, H.; Tang, J. Effects of citric acid and Lactobacillus plantarum on silage quality and bacterial diversity of king grass silage. Front. Microbiol, 1096. [Google Scholar] [CrossRef]
- Barboza, N.; Brenes-Guillen, L.; Uribe, L.; WingChing-Jones, R. Silage quality and bacterial diversity of silage inoculated with Listeria monocytogenes and Lacticaseibacillus paracasei_6714. Rev. Biol. Trop. 2023, 71, 50692. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef]
- Borges, E.B.; Araújo, C.A.; Monteiro, B.S.; Silva, A.S.; Albuquerque, L.F.; de Araújo, G.L.; Campos, F.S.; Gois, G.C.; de Souza, R.C.; de Araújo, A.O. Buffel grass pre-dried as a modulator of the fermentation, nutritional and aerobic stability profile of cactus pear silage. N. Z. J. Agric. 2023. [Google Scholar] [CrossRef]
- Amorim, D.S.; Edvan, R.L.; do Nascimento, R.R.; Bezerra, L.R.; de Araújo, M.J.; da Silva, A.L.; Meilezrski, F.; Nascimento, K.S. Fermentation profile and nutritional value of sesame silage compared to usual silages. Ital. J. Anim. Sci. 2020, 2020. 19, 19,230–239. [Google Scholar] [CrossRef]
- Dunière, L.; Xu, S.; Long, J.; Elekwachi, C.; Wang, Y.; Turkington, K.; Forster, R.; McAllister, T.A. Bacterial and fungal core microbiomes associated with small grain silages during ensiling and aerobic spoilage. BMC Microbiol. 2017, 2017. 17, 50. [Google Scholar] [CrossRef]
- Ennahar, S.; Cai, Y.M.; Fujita, Y. Phylogenic diversity of lactic acid bacteria associated with paddy rice silage as determined by 16S ribosomal DNA analysis. Appl. Environ. Microbiol. 2003, 2003. 69, 444–451. [Google Scholar] [CrossRef]
- Jaipolsaen, N.; Sangsritavong, S.; Uengwetwanit, T.; Angthong, P.; Plengviidhya, V.; Rungrassamee, W.; Yammuenart, S. Comparison of the effects of microbial inoculants on fermentation quality and microbiota in napier grass (Pennisetum purpureum) and corn (Zea mays) silage. Front. Microbiol, /: https, 4535. [Google Scholar]
- Dong, J.; Li, S.; Chen, X. ; Sun. Z.; Sun, Y.; Zhen, Y.; Qin, G.; Wang, T.; Demelash, N.; Zhang, X. Effects of Lactiplantibacillus plantarum inoculation on the quality and bacterial community of whole-crop corn silage at different harvest stages. Chem. Biol. Technol. Agric. [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biotechnol. [CrossRef]
- Dong, Z.; Lim, J.; Chen, L.; Wang, S.; Shao, T. Effects of freeze-thaw event on microbial community dynamics during red clover ensiling. Front. Microbiol. 2019, 2019. 10, 1559. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, Z.; Sa, D.; Hou, M.; Ge, G.; Wang, Z.; Jia, Y. The Potential Effects on Microbiota and Silage Fermentation of Alfalfa under Salt Stress. Front. Microbiol, 6886. [Google Scholar] [CrossRef]
- Sa, D.W.; Lu, Q.; Wang, Z.; Ge, G.; Sun, L.; Jia, Y. The potential and effects of salin-alkali alfalfa microbiota under salt stress on the fermentation quality and microbial. BMC Microbiol. 2021, 21, 149. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Che, C.; Zhong, Y.; Bu, X.; Huang, S.; Tahir, M.; Du, Z.; Liu, W.; Yang, W.; Li, J.; Wu, Y.; Zhang, Z.; Lian, J.; Xiao, Q.; Yan, Y. Effect of storage time on the silage quality and microbial community of mixed maize and faba bean in the Qinghai-Tibet plateau. Front. Microbiol, 0401. [Google Scholar] [CrossRef]
- Wang, W.; Tan, Z.; Gu, L.; Ma, H.; Wang, Z.; Wang, L.; Wu, G.; Qin, G.; Wang, Y.; Pang, H. Variation of microbial community and fermentation quality in corn silage treated with lactic bacteria and Artemisia argyi during aerobic exposure. Toxins 2022, 14, 349. [Google Scholar] [CrossRef]
- Ali, N.; Wang, S.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Microbial diversity and fermentation profile of red clover silage inoculated with reconstituted indigenous and exogenous epiphytic microbiota. Bioresour. Technol. 1236. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Wang, S.; Zhao, L.; Zhang, B.; Jia, W.; Zhai, Z.; Zhao, L.; Li, Y. Effects of antibacterial peptide-producing Bacillus subtilis, gallic acid, and cellulase on fermentation quality and bacterial community of whole-plant corn silage. Front. Microbiol. /: https, 3389. [Google Scholar]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 2013. 182, 1–15. [Google Scholar] [CrossRef]
- Tian, J.; Yin, X.; Zhang, J. Changes of the fermentation quality and microbial community during re-ensiling of sweet corn stalk silage. Ital. J. Anim. Sci, 1: 21; :1. [CrossRef]
- Huang, Y.; Liang, L.; Dai, S.; Wu, C.; Chen, C.; Hao, J. Effect of different regions and ensiling periods on fermentation quality and the bacterial community of whole-plant maize silage. Front. Microbiol. 2021, 3695. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O. C. M. : Ogunade, I. M.; Weinberg, Z.; Adesogan, A. T. Silage review: foodborne pathogens in silage and their mitigation by silage additives. J. Dairy Sci, 4: 101, 4132. [Google Scholar] [CrossRef]
- Lingaas, F.; Tveit, B. Etiology of acetonemia in Norwegian cattle. 2. Effect of butyric acid, valeric acid, and putrescine. J. Dairy Sci, 2: 75, 2433. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Si, Q.; Wang, Z.; Liu, W.; Liu, M.; Ge, G.; Jia, Y.; Du, S. Influence of cellulose or Lactiplantibacillus plantarum on the ensiling performance and bacterial community in mixed silage of alfalfa and Leymus chinensis. Microorganisms. 2023, 2023. 11, 426. [Google Scholar] [CrossRef]
Figure 1.
Temperature of maize silage treatments exposed to air over four days. T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract. D1 – D4 are the days of maize silage aerobic exposure.
Figure 1.
Temperature of maize silage treatments exposed to air over four days. T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract. D1 – D4 are the days of maize silage aerobic exposure.
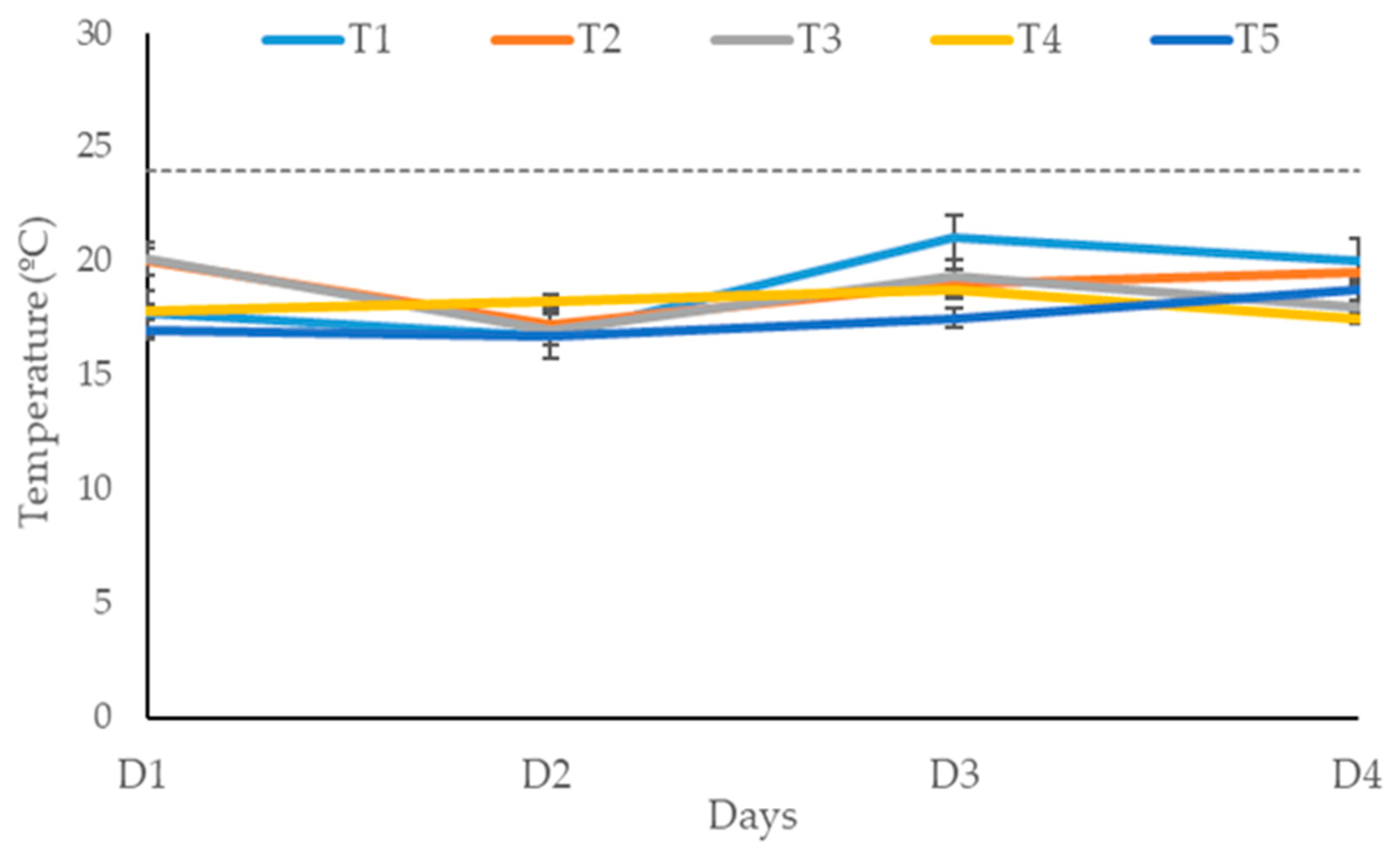
Figure 2.
Relative abundance of bacteria on maize silage at the phylum level (A) and genus level (B) as influenced by additives. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
Figure 2.
Relative abundance of bacteria on maize silage at the phylum level (A) and genus level (B) as influenced by additives. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
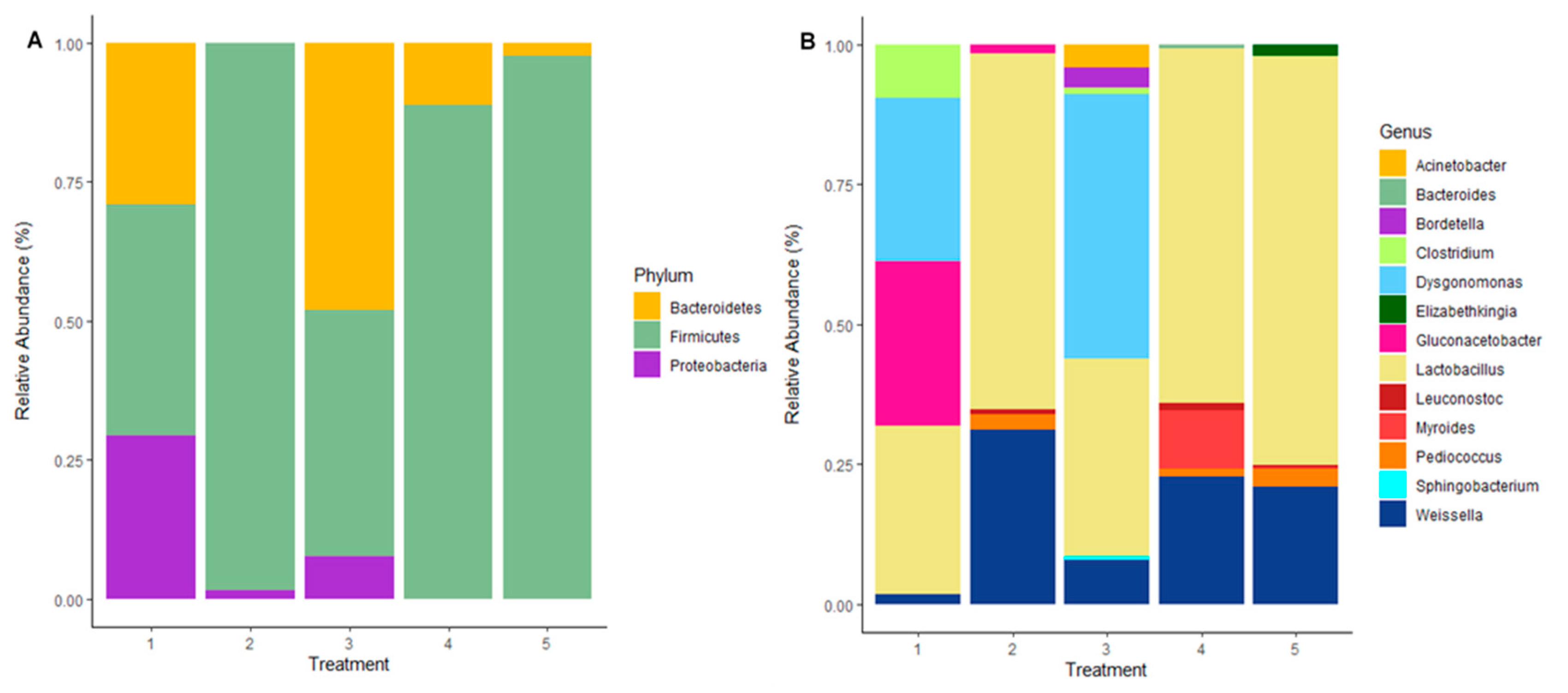
Figure 3.
Principal coordinate analysis (PCoA) of the bacterial community in maize silage depicting treatment differences. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
Figure 3.
Principal coordinate analysis (PCoA) of the bacterial community in maize silage depicting treatment differences. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
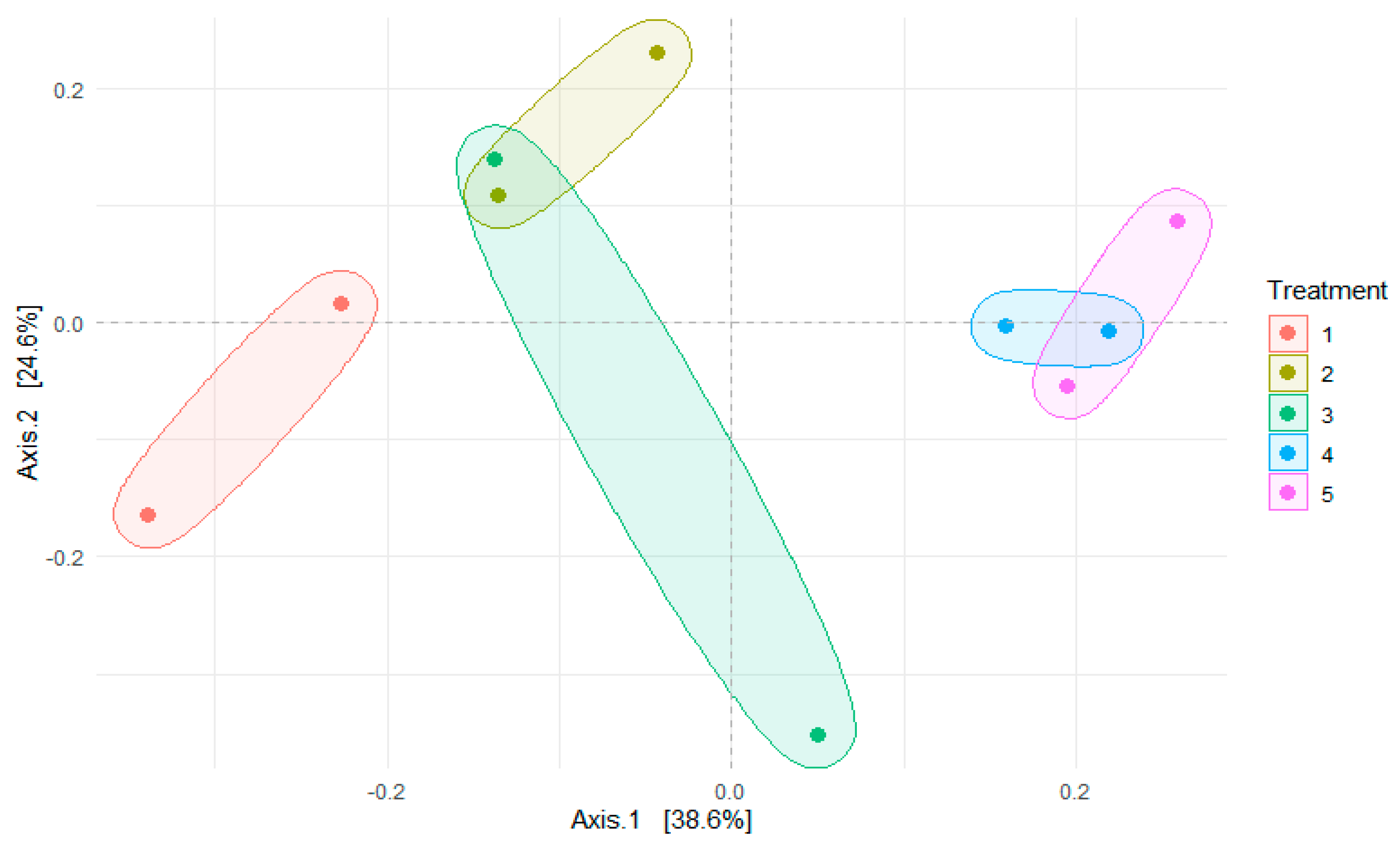
Figure 4.
Canonical correspondence analysis (CCA) shows the microbial composition relationships between maize silage treatments. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
Figure 4.
Canonical correspondence analysis (CCA) shows the microbial composition relationships between maize silage treatments. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).

Figure 5.
Analysis of dissimilarities of the bacterial composition of maize silages between treatments. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
Figure 5.
Analysis of dissimilarities of the bacterial composition of maize silages between treatments. Numbers 1, 2, 3, 4 and 5 denote treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
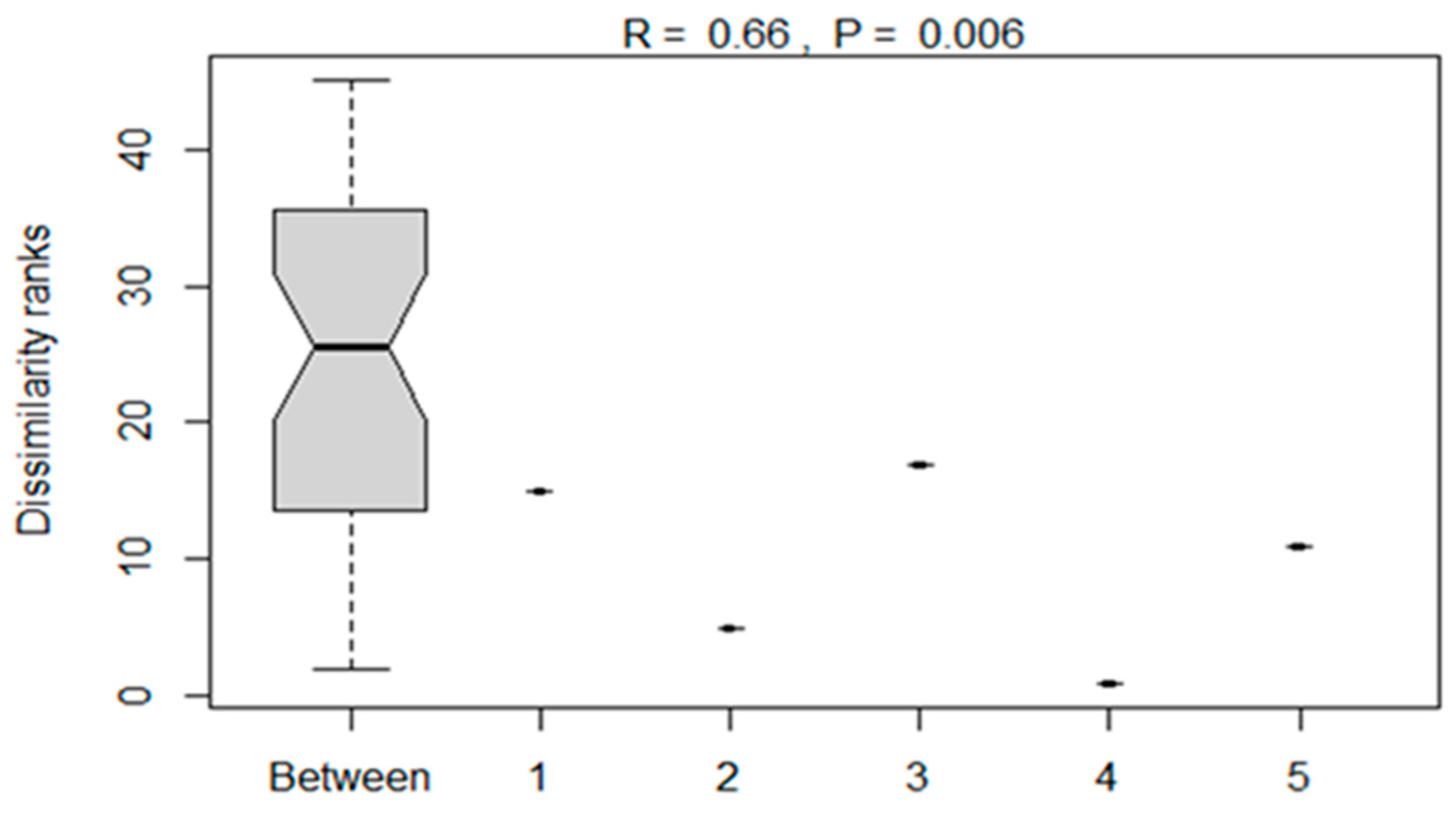
Figure 6.
Venn diagram to describe the common and unique OTUs among silage treatments. Numbers 1, 2, 3, 4 and 5 in bold denotes the treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).
Figure 6.
Venn diagram to describe the common and unique OTUs among silage treatments. Numbers 1, 2, 3, 4 and 5 in bold denotes the treatments (T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin extract; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin extract and T5, maize forage inoculated with LAB plus 3% tannin extract).


Table 1.
Effects of additives on fermentative quality of maize silage.
| Treatments | pH at day 0 | pH at day 75 | Silage DM% | DMR% | WL% |
|---|---|---|---|---|---|
| T1 | 5.13b | 3.52c | 31.1b | 82.4 | 7.0 |
| T2 | 5.15b | 3.53c | 33.3b | 93.2 | 2.2 |
| T3 | 5.16b | 3.55b | 32.7b | 89.3 | 3.9 |
| T4 | 5.17b | 3.52c | 36.4a | 93.8 | 2.3 |
| T5 | 5.29a | 3.60a | 36.5a | 87.2 | 4.8 |
| SEM | 0.017 | 0.005 | 0.825 | 2.63 | 1.13 |
| P-values | 0.0004 | <.0001 | 0.0032 | 0.0624 | 0.0674 |
DM, dry matter; DMR, dry matter recovery; WL, weight loss; T1, maize forage without LAB or tannin extract inoculant; T2, maize forage inoculated with LAB plus 1% tannin; T3, maize forage inoculated with LAB only; T4, maize forage inoculated with LAB plus 2% tannin and T5, maize forage inoculated with LAB plus 3% tannin. SEM, Standard Error of the mean. A,b,c letters in superscript within a column means different significance at p ≤ 0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.


Effects of Vachellia Mearnsii Tannin Extract as an Additive on Fermentation Quality, Aerobic Stability and Microbial modulation of Maize Silage
Thamsanqa Doctor Empire Mpanza
et al.
,
2023



MDPI Initiatives
Important Links
© 2024 MDPI (Basel, Switzerland) unless otherwise stated